

Evolutionary pathway to increased virulence and epidemic group A Streptococcus disease derived from 3,615 genome sequences
- PMID: 24733896
- PMCID: PMC4035937
- DOI: 10.1073/pnas.1403138111
Evolutionary pathway to increased virulence and epidemic group A Streptococcus disease derived from 3,615 genome sequences
Abstract
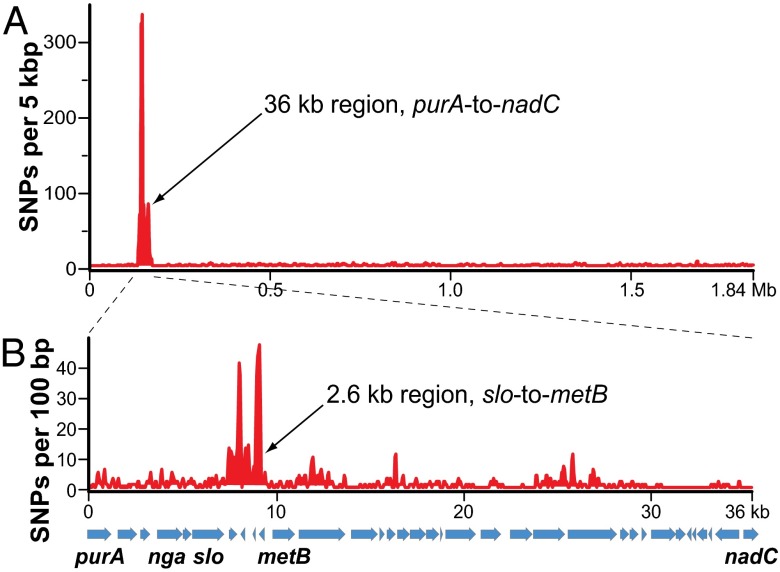
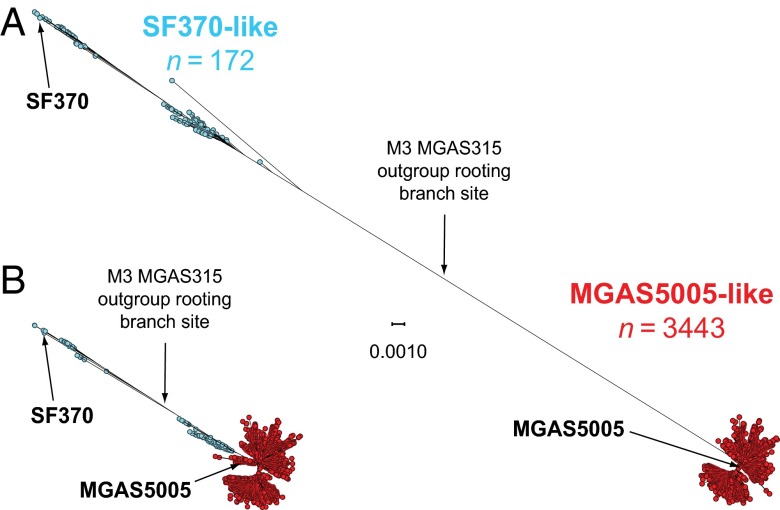
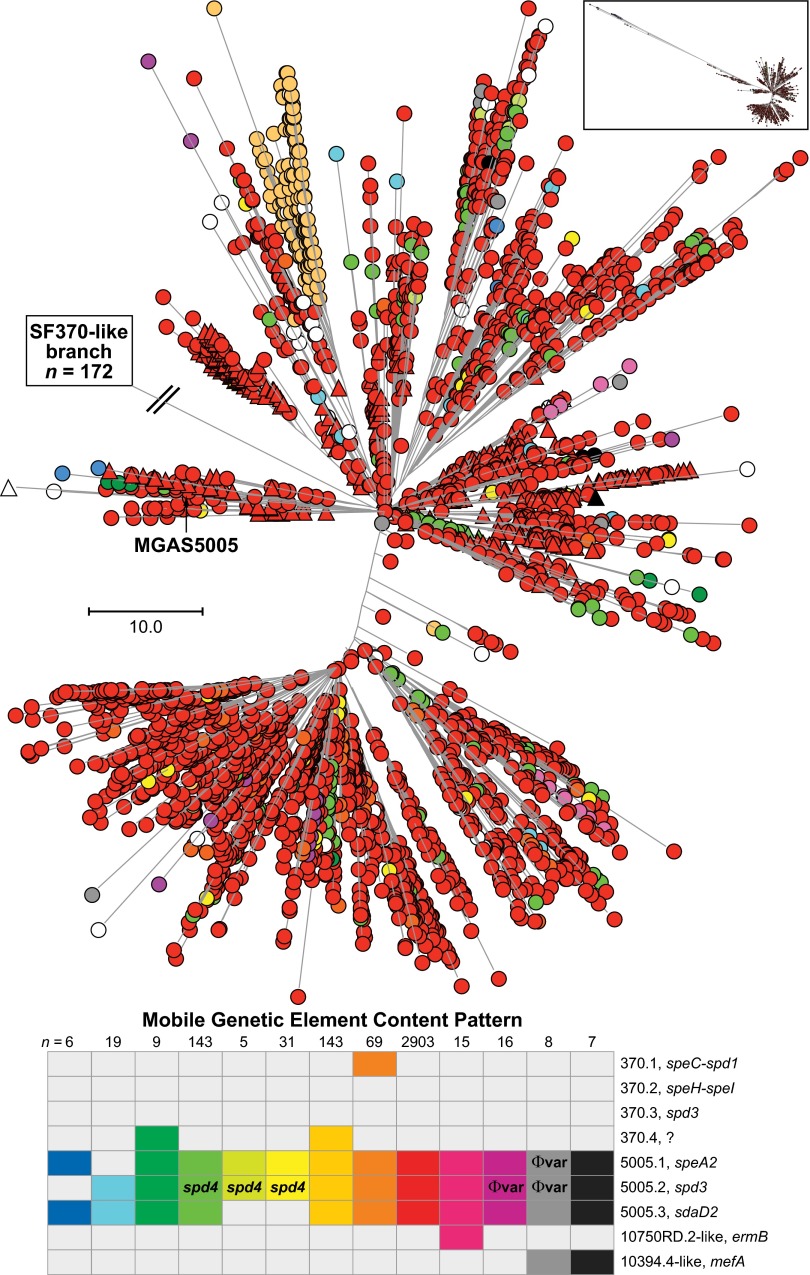
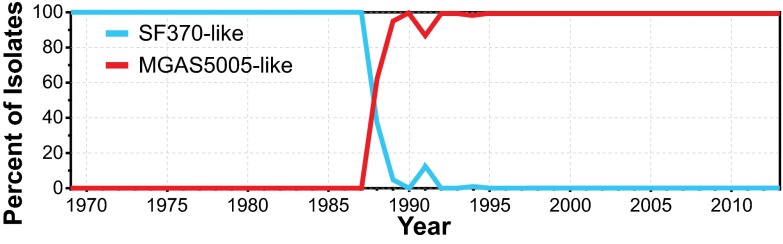
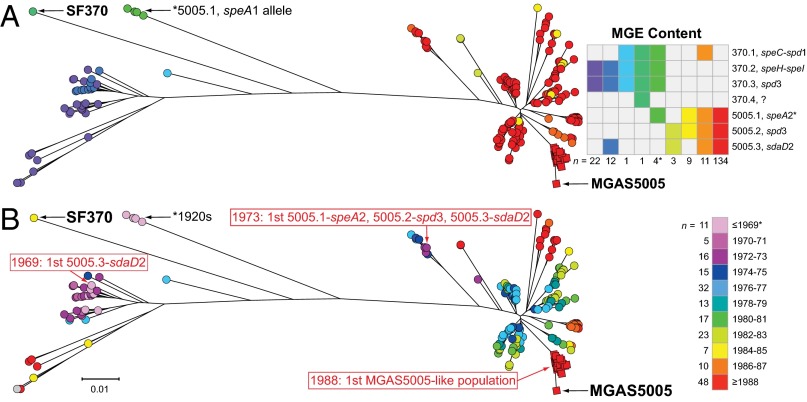
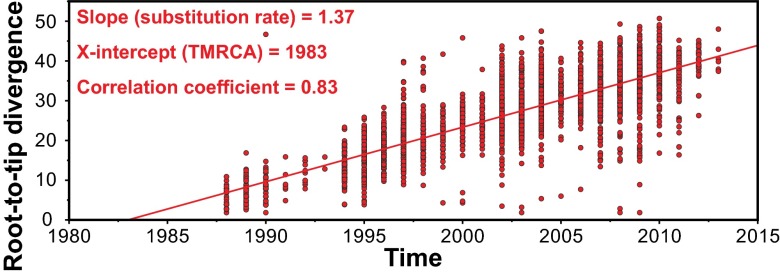
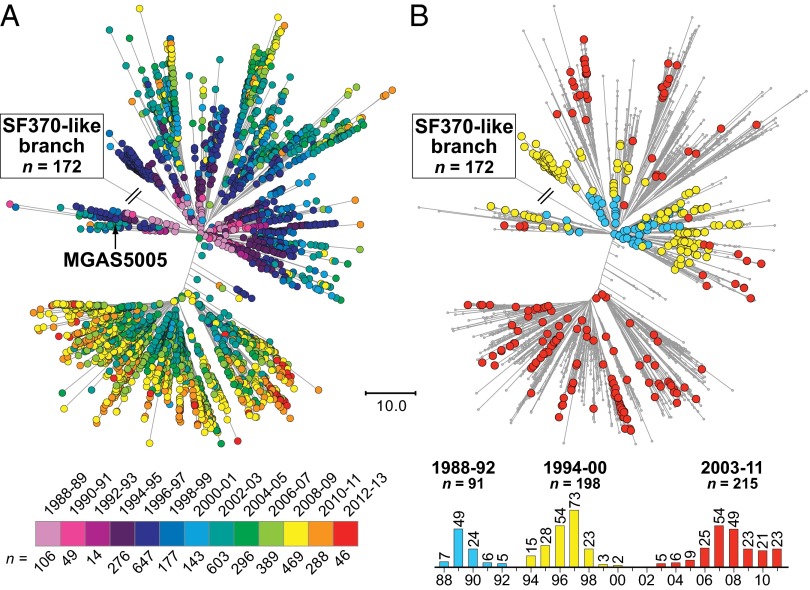
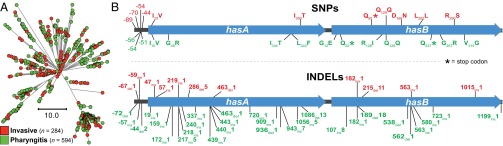
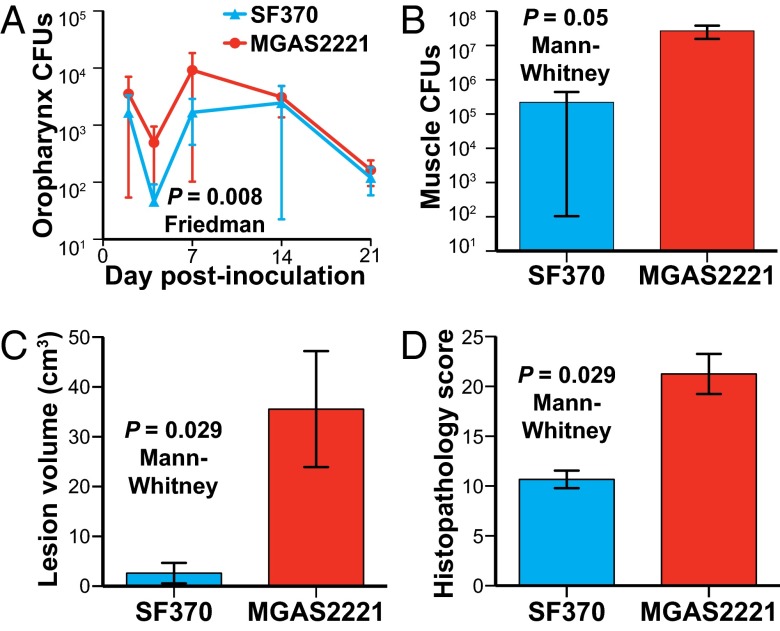
We sequenced the genomes of 3,615 strains of serotype Emm protein 1 (M1) group A Streptococcus to unravel the nature and timing of molecular events contributing to the emergence, dissemination, and genetic diversification of an unusually virulent clone that now causes epidemic human infections worldwide. We discovered that the contemporary epidemic clone emerged in stepwise fashion from a precursor cell that first contained the phage encoding an extracellular DNase virulence factor (streptococcal DNase D2, SdaD2) and subsequently acquired the phage encoding the SpeA1 variant of the streptococcal pyrogenic exotoxin A superantigen. The SpeA2 toxin variant evolved from SpeA1 by a single-nucleotide change in the M1 progenitor strain before acquisition by horizontal gene transfer of a large chromosomal region encoding secreted toxins NAD(+)-glycohydrolase and streptolysin O. Acquisition of this 36-kb region in the early 1980s into just one cell containing the phage-encoded sdaD2 and speA2 genes was the final major molecular event preceding the emergence and rapid intercontinental spread of the contemporary epidemic clone. Thus, we resolve a decades-old controversy about the type and sequence of genomic alterations that produced this explosive epidemic. Analysis of comprehensive, population-based contemporary invasive strains from seven countries identified strong patterns of temporal population structure. Compared with a preepidemic reference strain, the contemporary clone is significantly more virulent in nonhuman primate models of pharyngitis and necrotizing fasciitis. A key finding is that the molecular evolutionary events transpiring in just one bacterial cell ultimately have produced millions of human infections worldwide.
Keywords: flesh-eating disease; mobile genetic element; molecular clock; pathogenesis; phylogeography.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Comment in
-
Streptococcus pyogenes: the path to enhanced virulence.Clin Infect Dis. 2014 Jul 15;59(2):iv. Clin Infect Dis. 2014. PMID: 25105189 No abstract available.
References
-
- Bucher A, et al. Spectrum of disease in bacteraemic patients during a Streptococcus pyogenes serotype M-1 epidemic in Norway in 1988. Eur J Clin Microbiol Infect Dis. 1992;11(5):416–426. - PubMed
-
- Carapetis J, Robins-Browne R, Martin D, Shelby-James T, Hogg G. Increasing severity of invasive group A streptococcal disease in Australia: Clinical and molecular epidemiological features and identification of a new virulent M-nontypeable clone. Clin Infect Dis. 1995;21(5):1220–1227. - PubMed
-
- Carapetis JR, Steer AC, Mulholland EK, Weber M. The global burden of group A streptococcal diseases. Lancet Infect Dis. 2005;5(11):685–694. - PubMed
-
- Chelsom J, Halstensen A, Haga T, Høiby EA. Necrotising fasciitis due to group A streptococci in western Norway: Incidence and clinical features. Lancet. 1994;344(8930):1111–1115. - PubMed
Publication types
MeSH terms
Associated data
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
