

Regulation of mucosal immunity in the female reproductive tract: the role of sex hormones in immune protection against sexually transmitted pathogens
- PMID: 24734774
- PMCID: PMC4351777
- DOI: 10.1111/aji.12252
Regulation of mucosal immunity in the female reproductive tract: the role of sex hormones in immune protection against sexually transmitted pathogens
Abstract
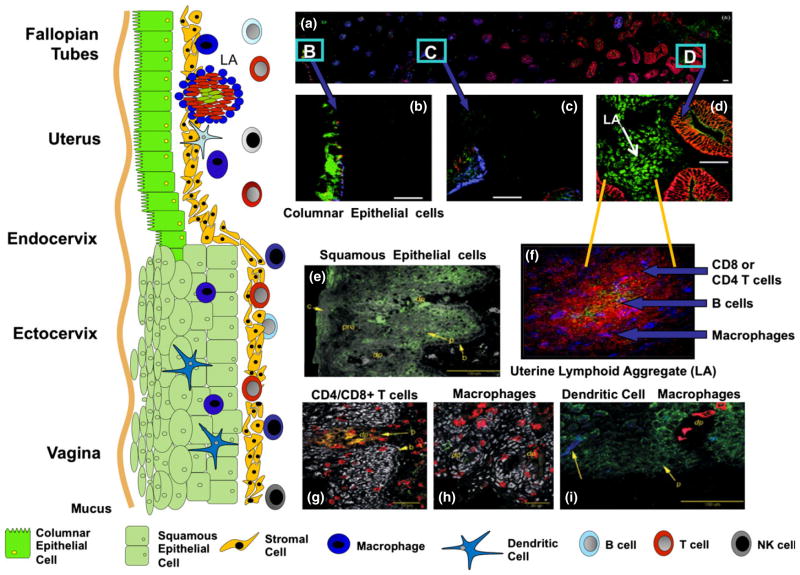
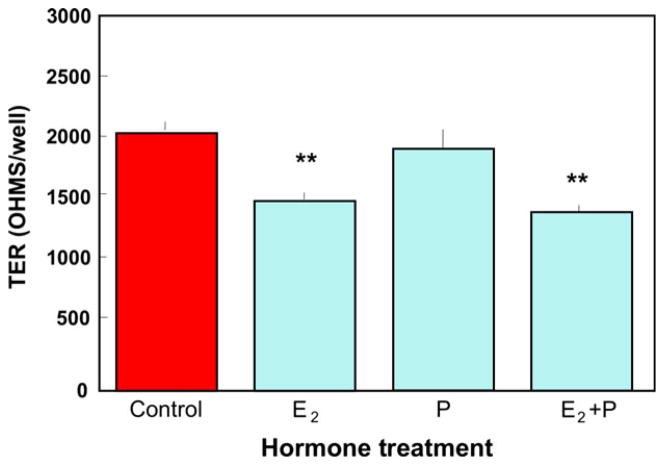
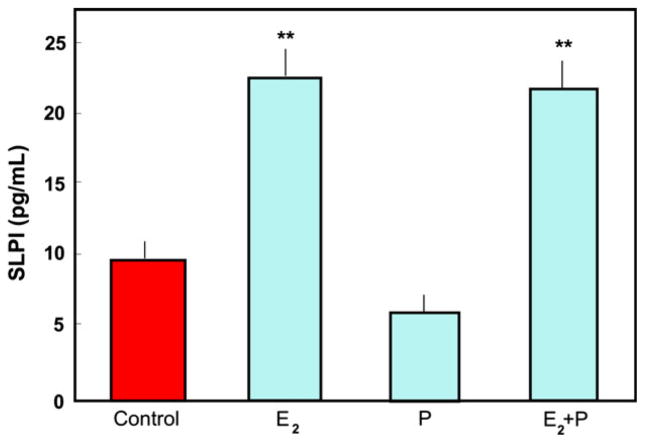
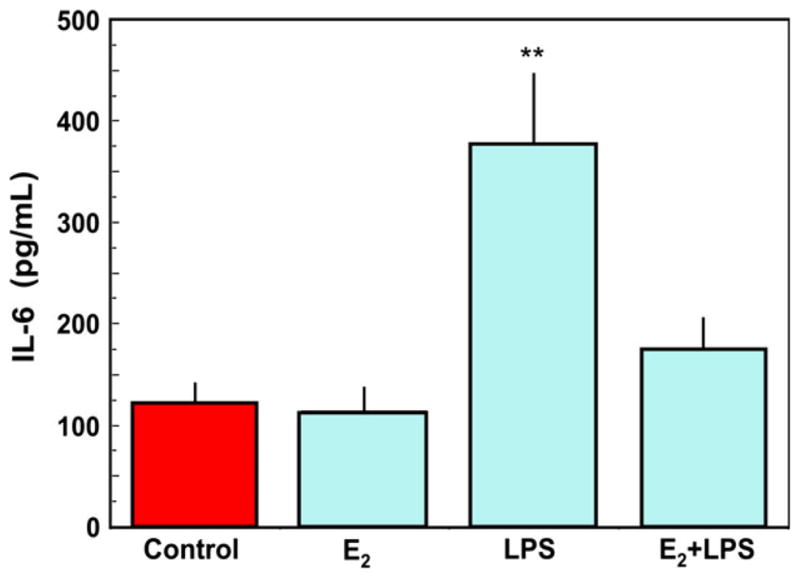
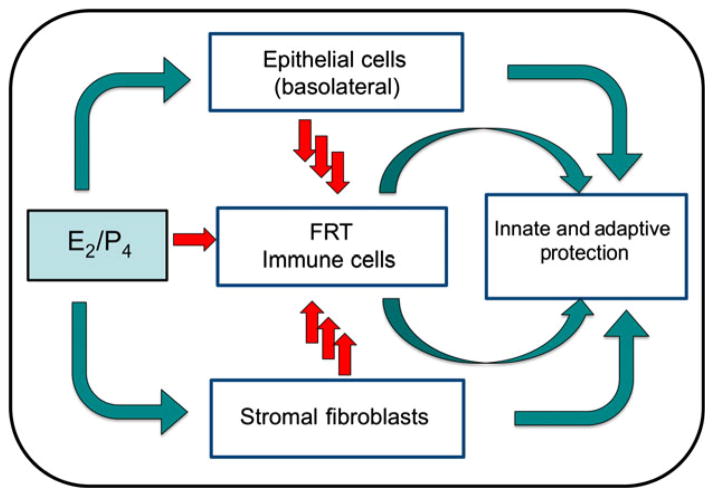
The immune system in the female reproductive tract (FRT) does not mount an attack against human immunodeficiency virus (HIV) or other sexually transmitted infections (STI) with a single endogenously produced microbicide or with a single arm of the immune system. Instead, the body deploys dozens of innate antimicrobials to the secretions of the FRT. Working together, these antimicrobials along with mucosal antibodies attack viral, bacterial, and fungal targets. Within the FRT, the unique challenges of protection against sexually transmitted pathogens coupled with the need to sustain the development of an allogeneic fetus, has evolved in such a way that sex hormones precisely regulate immune function to accomplish both tasks. The studies presented in this review demonstrate that estradiol (E2 ) and progesterone secreted during the menstrual cycle act both directly and indirectly on epithelial cells, fibroblasts and immune cells in the reproductive tract to modify immune function in a way that is unique to specific sites throughout the FRT. As presented in this review, studies from our laboratory and others demonstrate that the innate and adaptive immune systems are under hormonal control, that protection varies with the stage of the menstrual cycle and as such, is dampened during the secretory stage of the cycle to optimize conditions for fertilization and pregnancy. In doing so, a window of STI vulnerability is created during which potential pathogens including HIV enter the reproductive tract to infect host targets.
Keywords: Epithelial cells; female reproductive tract; fibroblasts; immune cells; mucosal immunity; sex hormones.
© 2014 John Wiley & Sons A/S. Published by John Wiley & Sons Ltd.
Figures
References
-
- UNAIDS. AIDS Epidemic Update, Joint United Nations Programme on HIV/AIDS (UNAIDS) and World Health Organization (WHO) Geneva, Switzerland: 2008. UNAIDS 2008.
-
- UNAIDS. AIDS Epidemic Update JUNPoHAUaWHOW. Geneva, Switzerland: 2007.
-
- Ogra P, Yamanaka T, Losonsky GA. Reproductive Immunology. In: Fleicher N, editor. Local immunologic defenses in the genital tract. New York: Alan R. Liss, Inc; 1981. pp. 381–394. - PubMed
-
- Wira CR, Fahey JV, White HD, Yeaman GR, Given AL, Howell AL. The mucosal immune system in the human female reproductive tract: influence of stage of the menstrual cycle and menopause on mucosal immunity in the uterus. In: Glasser S, Aplin J, Guidice L, Tabibzadeh S, editors. The Endometrium. New York: Taylor and Francis; 2002. pp. 371–404.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
