

Trade-offs in engineering sugar utilization pathways for titratable control
- PMID: 24735079
- PMCID: PMC4384834
- DOI: 10.1021/sb400162z
Trade-offs in engineering sugar utilization pathways for titratable control
Abstract
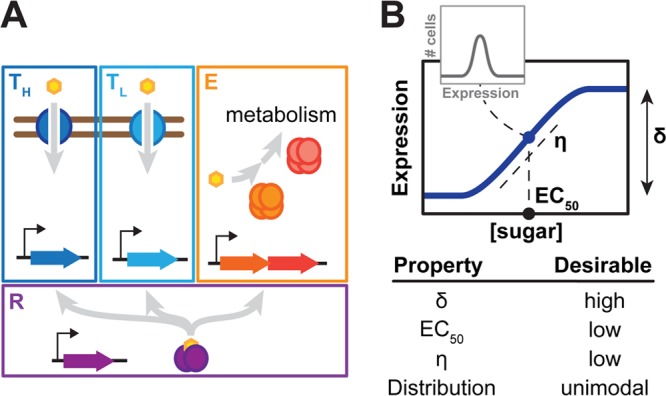
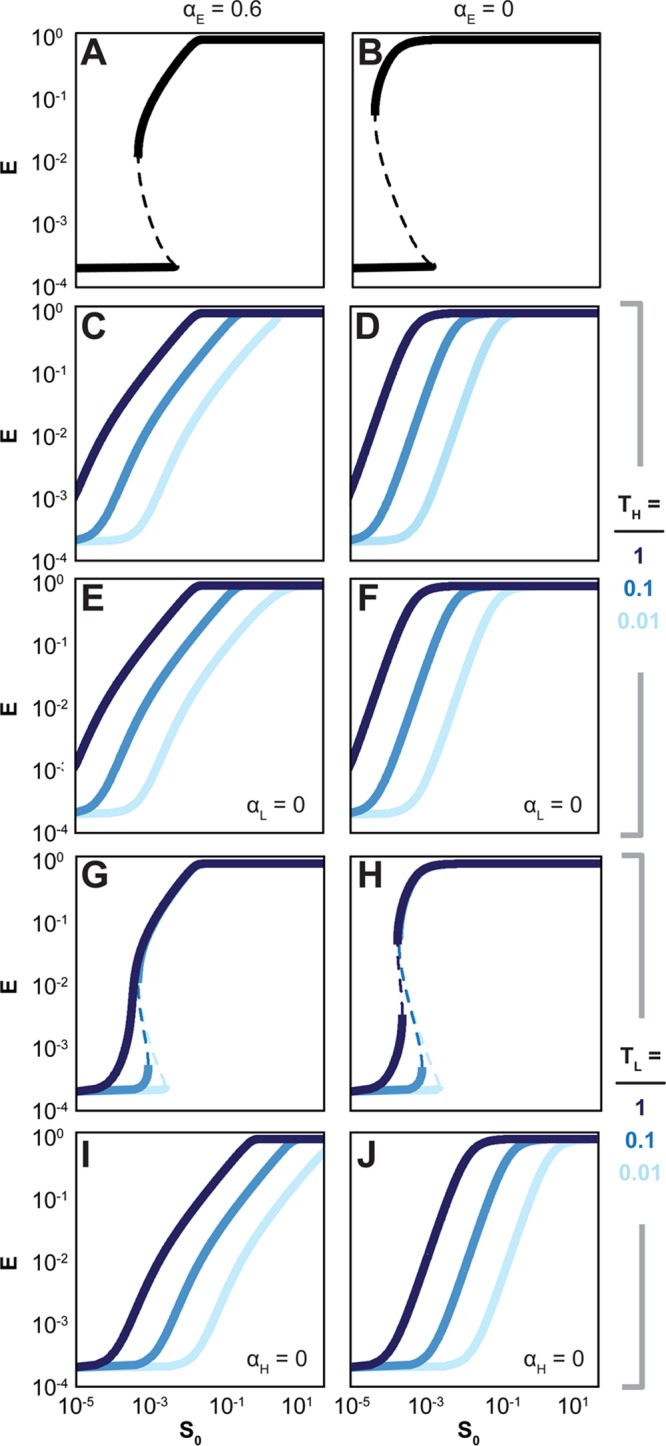
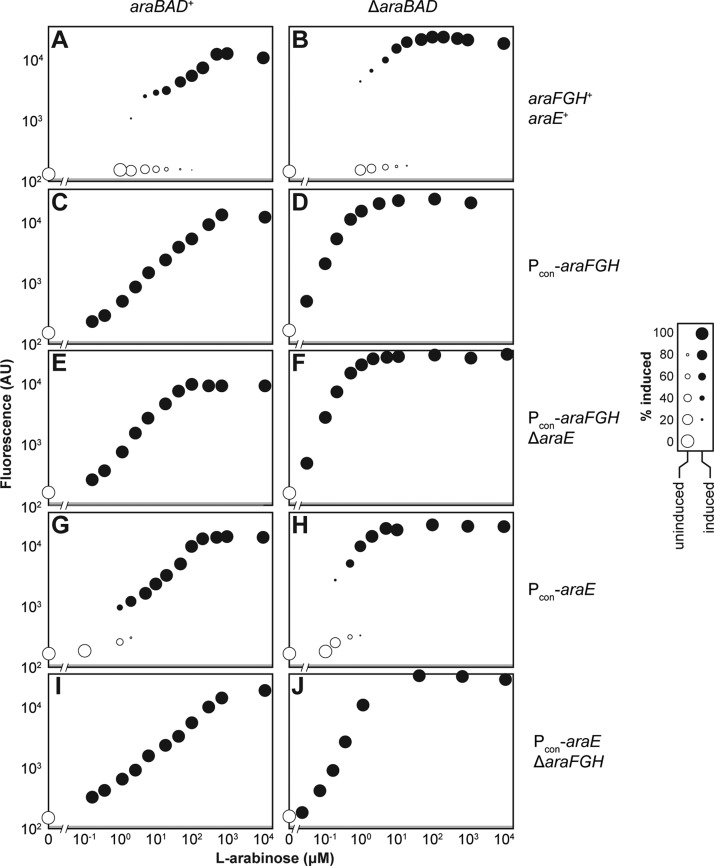
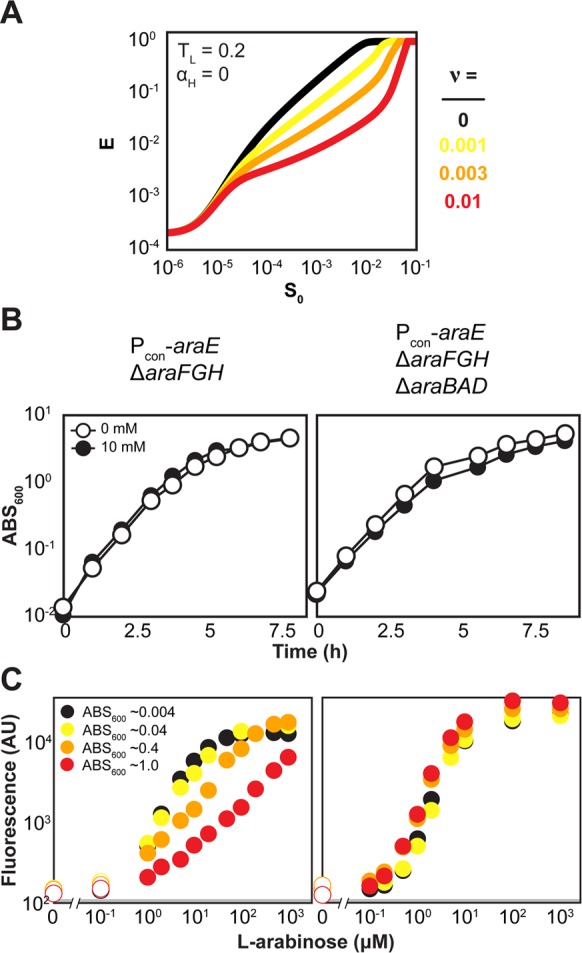
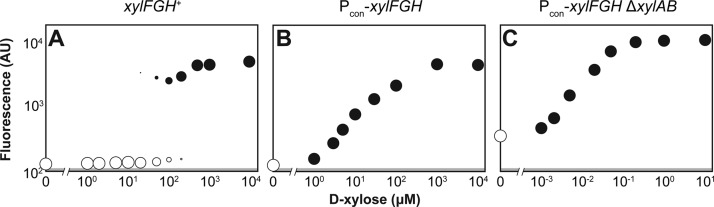
Titratable systems are common tools in metabolic engineering to tune the levels of enzymes and cellular components as part of pathway optimization. For nonmodel microorganisms with limited genetic tools, inducible sugar utilization pathways offer built-in titratable systems. However, these pathways can exhibit undesirable single-cell behaviors that hamper the uniform and tunable control of gene expression. Here, we applied mathematical modeling and single-cell measurements of L-arabinose utilization in Escherichia coli to systematically explore how sugar utilization pathways can be altered to achieve desirable inducible properties. We found that different pathway alterations, such as the removal of catabolism, constitutive expression of high-affinity or low-affinity transporters, or further deletion of the other transporters, came with trade-offs specific to each alteration. For instance, sugar catabolism improved the uniformity and linearity of the response at the cost of requiring higher sugar concentrations to induce the pathway. Within these alterations, we also found that a uniform and linear response could be achieved with a single alteration: constitutively expressing the high-affinity transporter. Equivalent modifications to the D-xylose utilization pathway yielded similar responses, demonstrating the applicability of our observations. Overall, our findings indicate that there is no ideal set of typical alterations when co-opting natural utilization pathways for titratable control and suggest design rules for manipulating these pathways to advance basic genetic studies and the metabolic engineering of microorganisms for optimized chemical production.
Keywords: Escherichia coli; carbon catabolism; feedback; inducible systems; linearization; systems biology.
Figures
References
-
- Hawkins K. M.; Smolke C. D. (2006) The regulatory roles of the galactose permease and kinase in the induction response of the GAL network in Saccharomyces cerevisiae. J. Biol. Chem. 281, 13485–13492. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
