

DNA methylation dynamics during epigenetic reprogramming in the germline and preimplantation embryos
- PMID: 24736841
- PMCID: PMC4003274
- DOI: 10.1101/gad.234294.113
DNA methylation dynamics during epigenetic reprogramming in the germline and preimplantation embryos
Abstract
Methylation of DNA is an essential epigenetic control mechanism in mammals. During embryonic development, cells are directed toward their future lineages, and DNA methylation poses a fundamental epigenetic barrier that guides and restricts differentiation and prevents regression into an undifferentiated state. DNA methylation also plays an important role in sex chromosome dosage compensation, the repression of retrotransposons that threaten genome integrity, the maintenance of genome stability, and the coordinated expression of imprinted genes. However, DNA methylation marks must be globally removed to allow for sexual reproduction and the adoption of the specialized, hypomethylated epigenome of the primordial germ cell and the preimplantation embryo. Recent technological advances in genome-wide DNA methylation analysis and the functional description of novel enzymatic DNA demethylation pathways have provided significant insights into the molecular processes that prepare the mammalian embryo for normal development.
Keywords: DNA methylation; development; epigenetics; reprogramming.
Figures
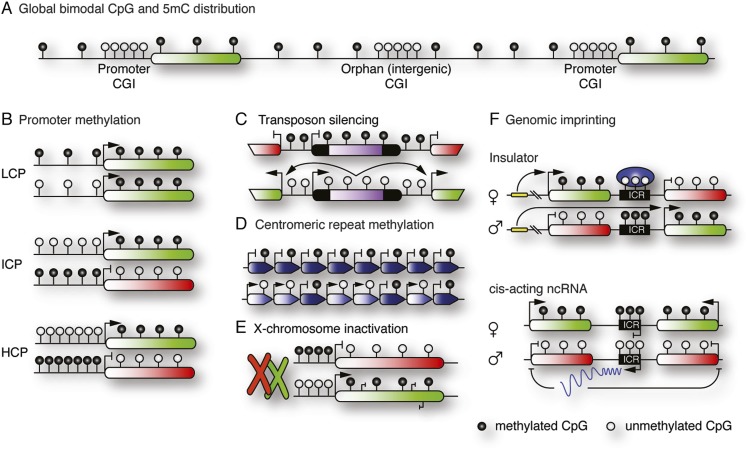
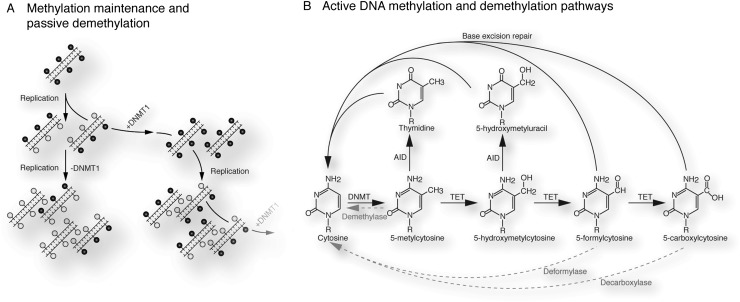
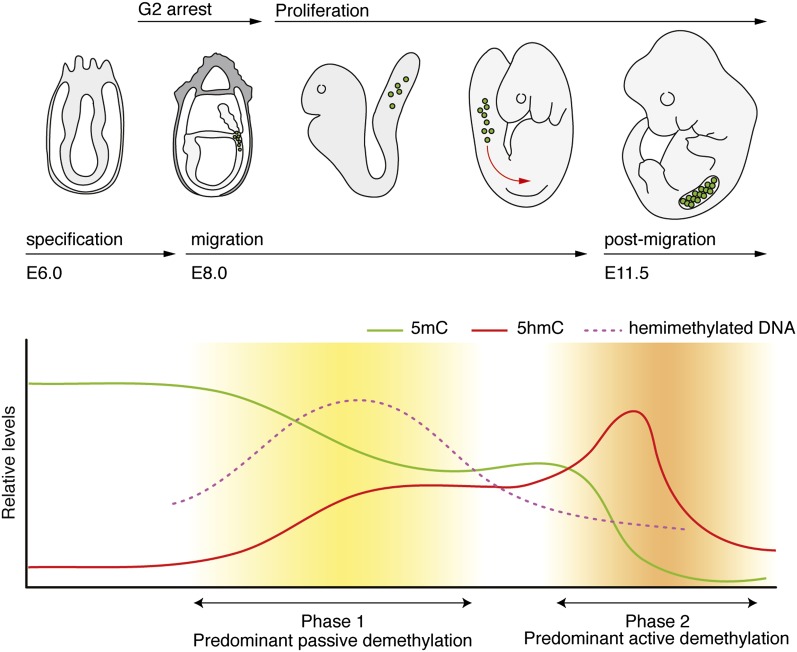
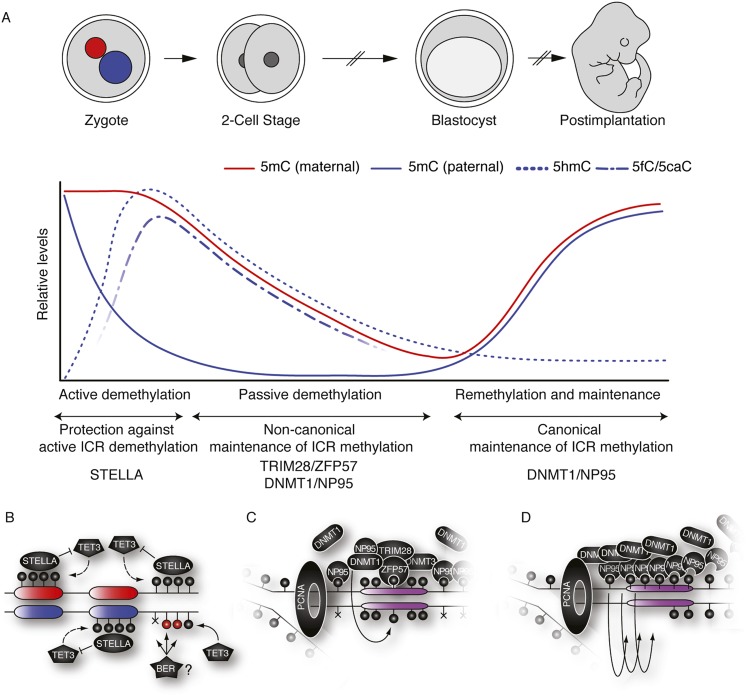
References
-
- Abdalla H, Yoshizawa Y, Hochi S 2009. Active demethylation of paternal genome in mammalian zygotes. J Reprod Dev 55: 356–360 - PubMed
-
- Anastasiadou C, Malousi A, Maglaveras N, Kouidou S 2011. Human epigenome data reveal increased CpG methylation in alternatively spliced sites and putative exonic splicing enhancers. DNA Cell Biol 30: 267–275 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources