

Mechanism of inhibition of the GluA1 AMPA receptor channel opening by the 2,3-benzodiazepine compound GYKI 52466 and a N-methyl-carbamoyl derivative
- PMID: 24738995
- PMCID: PMC4025570
- DOI: 10.1021/bi5002079
Mechanism of inhibition of the GluA1 AMPA receptor channel opening by the 2,3-benzodiazepine compound GYKI 52466 and a N-methyl-carbamoyl derivative
Abstract
2,3-Benzodiazepine derivatives, also known as GYKI compounds, represent a group of the most promising synthetic inhibitors of α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid (AMPA) receptors. Here we investigate the mechanism of inhibition of the GluA1 channel opening and the site of inhibition by GYKI 52466 and its N-3 methyl-carbamoyl derivative, which we term as BDZ-f. GluA1 is a key AMPA receptor subunit involved in the brain function. Excessive activity and elevated expression of GluA1, however, has been implicated in a number of neurological disorders. Using a laser-pulse photolysis technique, which provides ∼60 μs resolution, we measured the effect of these inhibitors on the rate of GluA1 channel opening and the amplitude of the glutamate-induced whole-cell current. We found that both compounds inhibit GluA1 channel noncompetitively. Addition of an N-3 methyl-carbamoyl group to the diazepine ring with the azomethine feature (i.e., GYKI 52466) improves the potency of the resulting compound or BDZ-f without changing the site of binding. This site, which we previously termed as the "M" site on the GluA2 AMPA receptor subunit, therefore favorably accommodates an N-3 acylating group. On the basis of the magnitude of the inhibition constants for the same inhibitors but different receptors, the "M" sites on GluA1 and GuA2 are different. Overall, the "M" site or the binding environment on GluA2 accommodates the same compounds better, or the same inhibitors show stronger potency on GluA2, as we have reported previously [ Wang et al. Biochemistry ( 2011 ) 50 , 7284 - 7293 ]. However, acylating the N-3 position to occupy the N-3 side pocket of the "M" site can significantly narrow the difference and improve the potency of a resulting compound on GluA1.
Figures


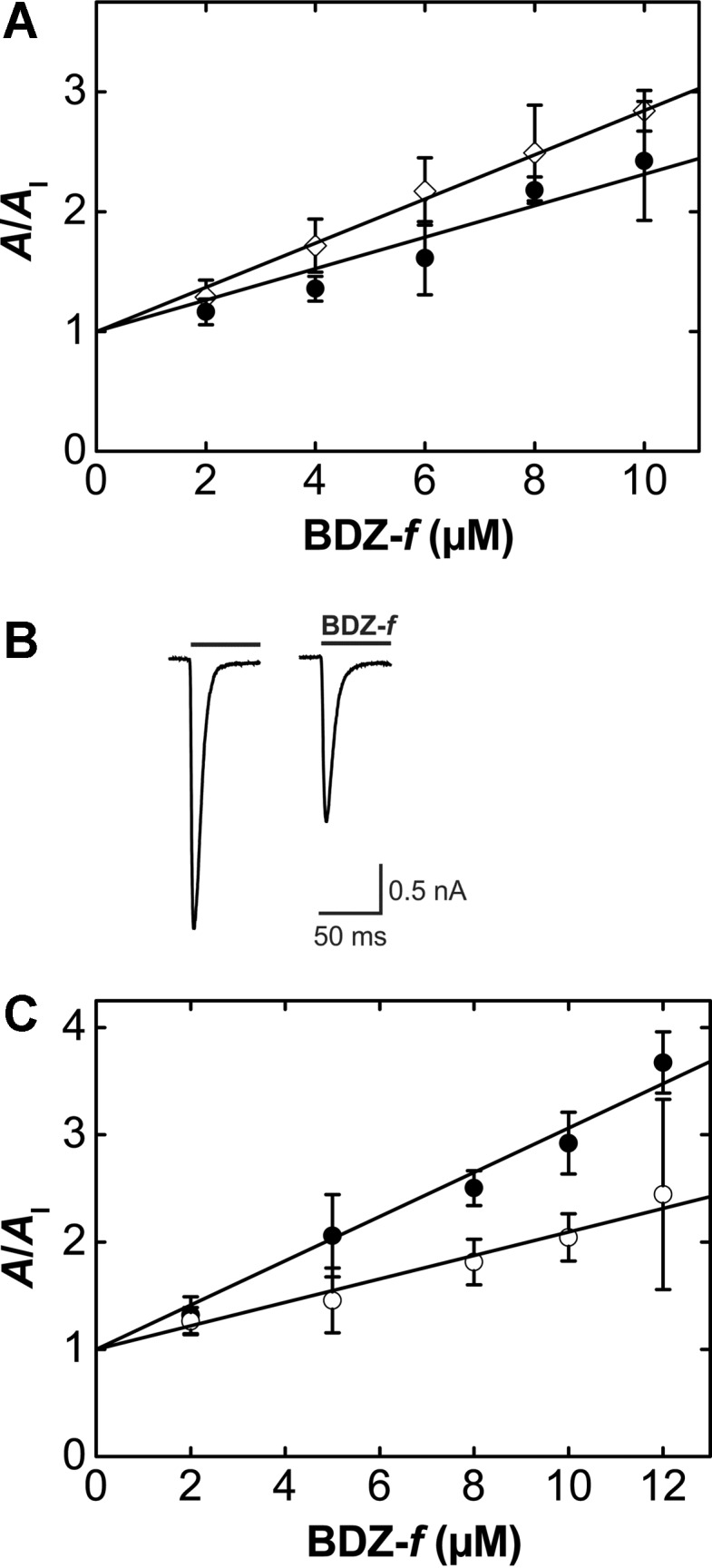
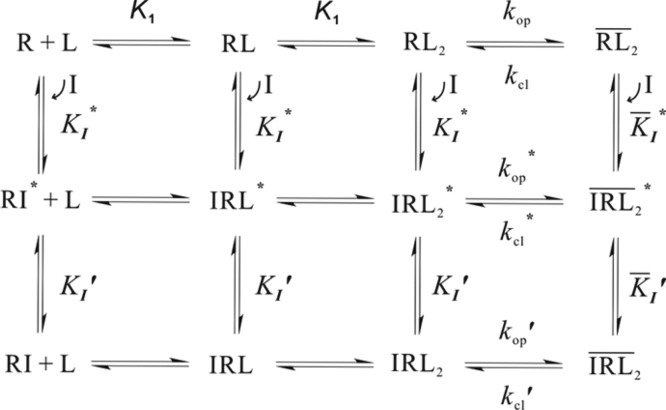
 2 refer to the open-channel state.
2 refer to the open-channel state.

Similar articles
-
Mechanism of inhibition of the GluA2 AMPA receptor channel opening: consequences of adding an N-3 methylcarbamoyl group to the diazepine ring of 2,3-benzodiazepine derivatives.Biochemistry. 2011 Aug 23;50(33):7284-93. doi: 10.1021/bi2007977. Epub 2011 Jul 28. Biochemistry. 2011. PMID: 21751782 Free PMC article.
-
Mechanism of inhibition of the GluR2 AMPA receptor channel opening by 2,3-benzodiazepine derivatives.Biochemistry. 2008 Jan 22;47(3):1061-9. doi: 10.1021/bi700782x. Epub 2007 Dec 28. Biochemistry. 2008. PMID: 18161947
-
Mechanism of inhibition of GluA2 AMPA receptor channel opening by 2,3-benzodiazepine derivatives: functional consequences of replacing a 7,8-methylenedioxy with a 7,8-ethylenedioxy moiety.Biochemistry. 2012 Feb 28;51(8):1787-95. doi: 10.1021/bi2017552. Epub 2012 Feb 13. Biochemistry. 2012. PMID: 22304561 Free PMC article.
-
Negative allosteric modulation of AMPA-preferring receptors by the selective isomer GYKI 53784 (LY303070), a specific non-competitive AMPA antagonist.CNS Drug Rev. 2002 Fall;8(3):235-54. doi: 10.1111/j.1527-3458.2002.tb00227.x. CNS Drug Rev. 2002. PMID: 12353057 Free PMC article. Review.
-
Non-competitive AMPA antagonists of 2,3-benzodiazepine type.Curr Pharm Des. 2002;8(10):913-39. doi: 10.2174/1381612024607081. Curr Pharm Des. 2002. PMID: 11945139 Review.
Cited by
-
Mechanism-based design of 2,3-benzodiazepine inhibitors for AMPA receptors.Acta Pharm Sin B. 2015 Nov;5(6):500-5. doi: 10.1016/j.apsb.2015.07.007. Epub 2015 Sep 26. Acta Pharm Sin B. 2015. PMID: 26713266 Free PMC article. Review.
-
Allosteric competition and inhibition in AMPA receptors.Nat Struct Mol Biol. 2024 Nov;31(11):1669-1679. doi: 10.1038/s41594-024-01328-0. Epub 2024 Jun 4. Nat Struct Mol Biol. 2024. PMID: 38834914 Free PMC article.
-
Allosteric Competition and Inhibition in AMPA Receptors.bioRxiv [Preprint]. 2023 Nov 28:2023.11.28.569057. doi: 10.1101/2023.11.28.569057. bioRxiv. 2023. Update in: Nat Struct Mol Biol. 2024 Nov;31(11):1669-1679. doi: 10.1038/s41594-024-01328-0. PMID: 38076818 Free PMC article. Updated. Preprint.
-
Noncompetitive antagonists induce cooperative AMPA receptor channel gating.J Gen Physiol. 2019 Feb 4;151(2):156-173. doi: 10.1085/jgp.201812209. Epub 2019 Jan 8. J Gen Physiol. 2019. PMID: 30622133 Free PMC article.
References
-
- Dingledine R.; Borges K.; Bowie D.; Traynelis S. F. (1999) The glutamate receptor ion channels. Pharmacol. Rev. 51, 7–61. - PubMed
-
- Squire L. R. (1992) Memory and the hippocampus: a synthesis from findings with rats, monkeys, and humans. Psychol. Rev. 99, 195–231. - PubMed
-
- Shi S.; Hayashi Y.; Esteban J. A.; Malinow R. (2001) Subunit-specific rules governing AMPA receptor trafficking to synapses in hippocampal pyramidal neurons. Cell 105, 331–343. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
