

Diabetes irreversibly depletes bone marrow-derived mesenchymal progenitor cell subpopulations
- PMID: 24740572
- PMCID: PMC4429348
- DOI: 10.2337/db13-1366
Diabetes irreversibly depletes bone marrow-derived mesenchymal progenitor cell subpopulations
Abstract
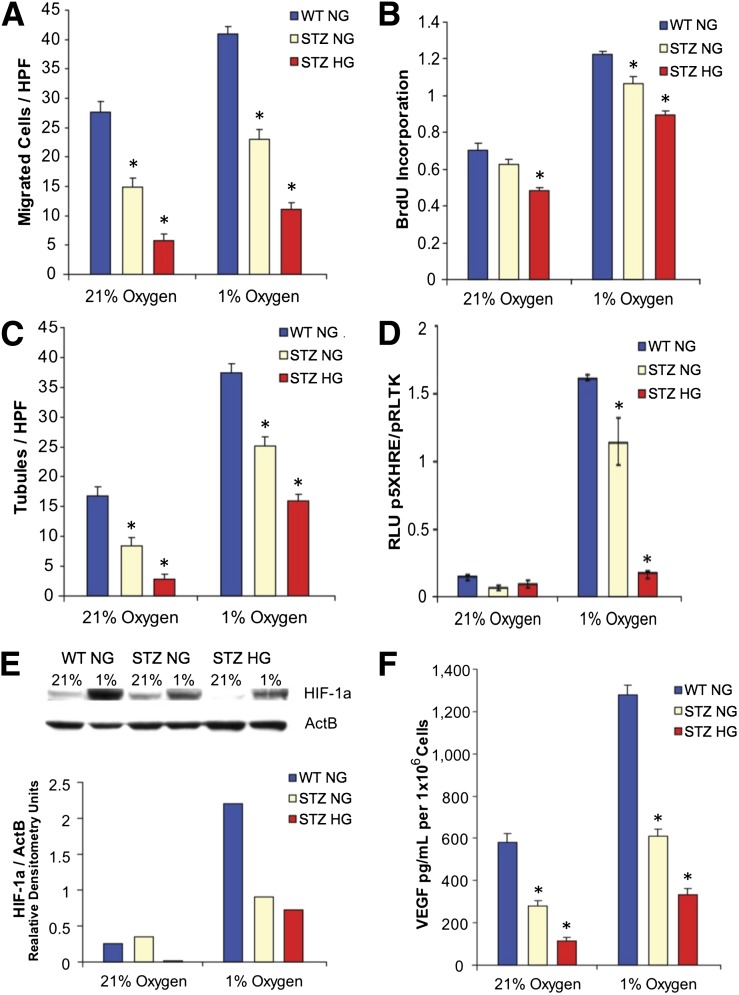
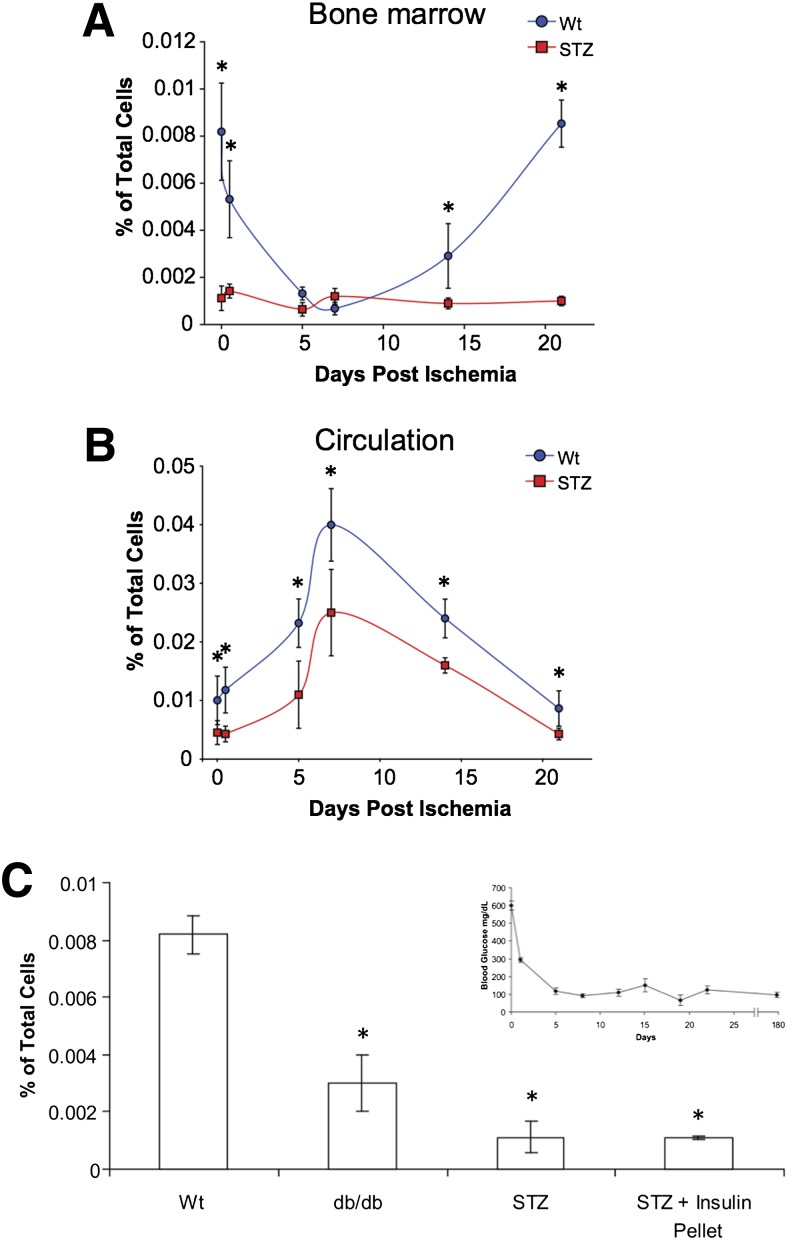
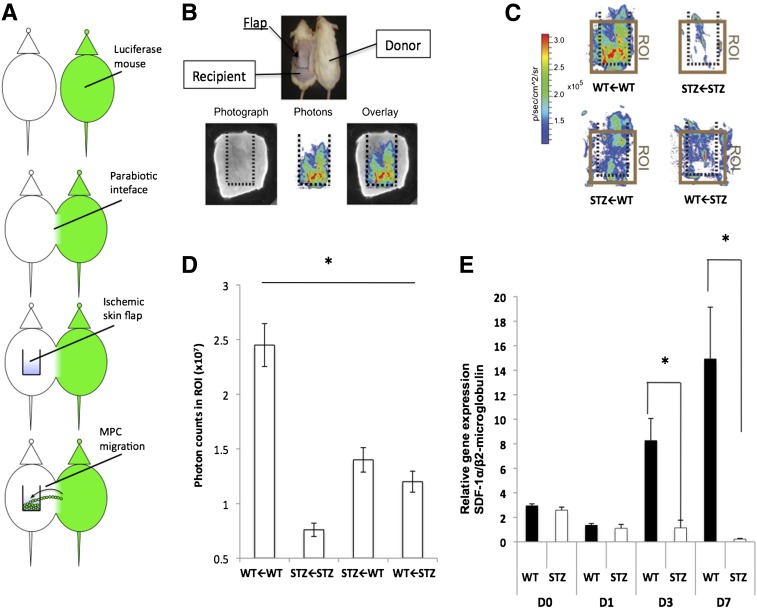
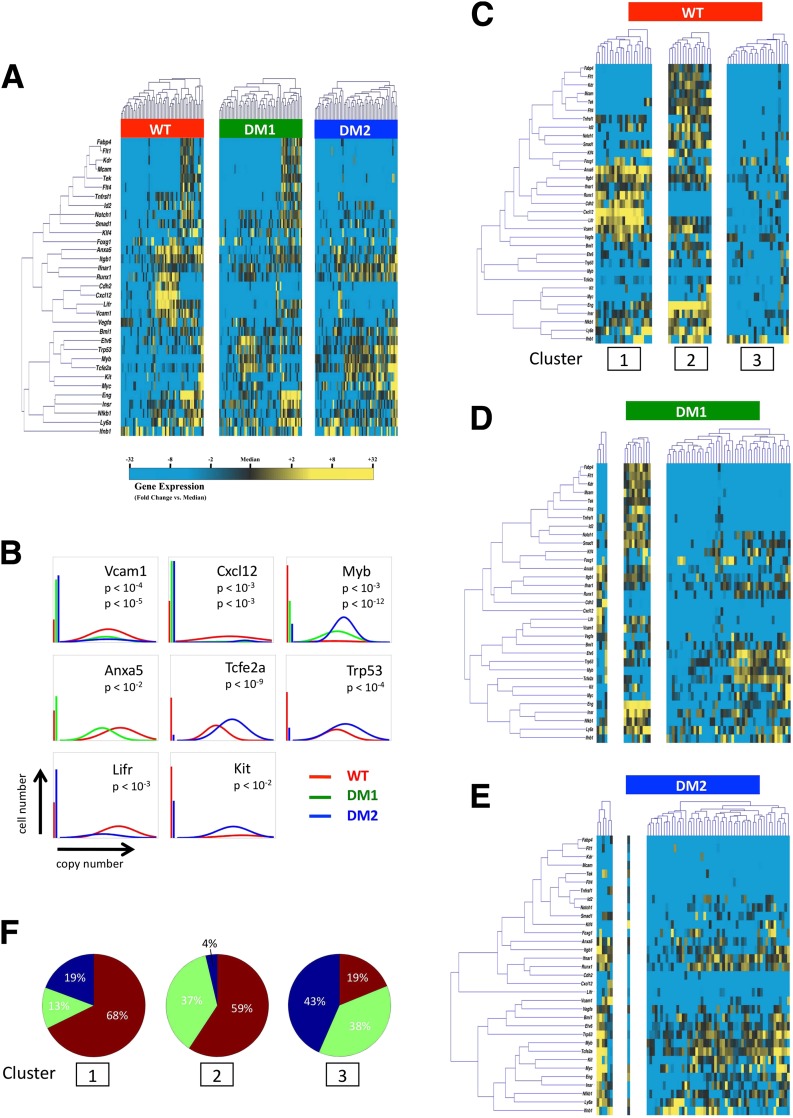
Diabetic vascular pathology is largely attributable to impairments in tissue recovery from hypoxia. Circulating progenitor cells have been postulated to play a role in ischemic recovery, and deficiencies in these cells have been well described in diabetic patients. Here, we examine bone marrow-derived mesenchymal progenitor cells (BM-MPCs) that have previously been shown to be important for new blood vessel formation and demonstrate significant deficits in the context of diabetes. Further, we determine that this dysfunction is attributable to intrinsic defects in diabetic BM-MPCs that are not correctable by restoring glucose homeostasis. We identify two transcriptionally distinct subpopulations that are selectively depleted by both type 1 and type 2 diabetes, and these subpopulations have provasculogenic expression profiles, suggesting that they are vascular progenitor cells. These results suggest that the clinically observed deficits in progenitor cells may be attributable to selective and irreversible depletion of progenitor cell subsets in patients with diabetes.
© 2014 by the American Diabetes Association. Readers may use this article as long as the work is properly cited, the use is educational and not for profit, and the work is not altered.
Figures
Comment in
-
Diabetes: Functional impairment of bone marrow progenitor cells in diabetes mellitus.Nat Rev Endocrinol. 2014 Jul;10(7):379. doi: 10.1038/nrendo.2014.66. Epub 2014 May 6. Nat Rev Endocrinol. 2014. PMID: 24798060 No abstract available.
References
-
- Zimmet P, Alberti KG, Shaw J. Global and societal implications of the diabetes epidemic. Nature 2001;414:782–787 - PubMed
-
- Annual number (in thousands) of new cases of diagnosed diabetes among adults aged 18–79 years, United States, 1980–2010 [article online], 2010. Available from http://www.cdc.gov/diabetes/statistics/incidence/fig1.htm. Accessed 14 June 2013
-
- Narayan KM, Boyle JP, Geiss LS, Saaddine JB, Thompson TJ. Impact of recent increase in incidence on future diabetes burden: U.S., 2005-2050. Diabetes Care 2006;29:2114–2116 - PubMed
-
- Caro JJ, Ward AJ, O'Brien JA. Lifetime costs of complications resulting from type 2 diabetes in the U.S. Diabetes Care 2002;25:476–481 - PubMed
-
- Fox CS, Coady S, Sorlie PD, et al. . Increasing cardiovascular disease burden due to diabetes mellitus: the Framingham Heart Study. Circulation 2007;115:1544–1550 - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
