

mTORC1-independent reduction of retinal protein synthesis in type 1 diabetes
- PMID: 24740573
- PMCID: PMC4141367
- DOI: 10.2337/db14-0235
mTORC1-independent reduction of retinal protein synthesis in type 1 diabetes
Abstract
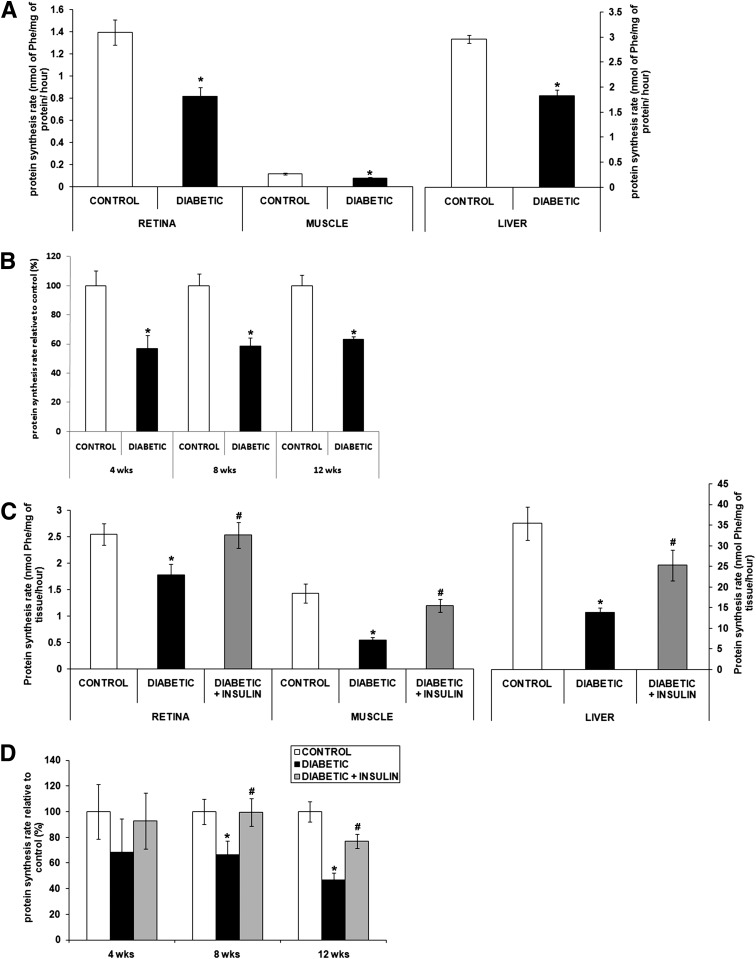
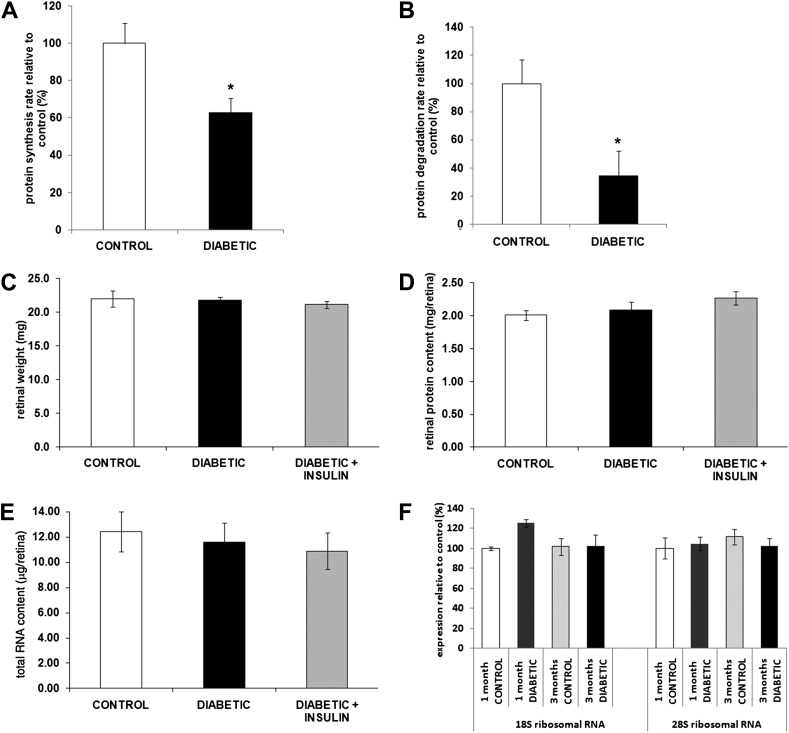
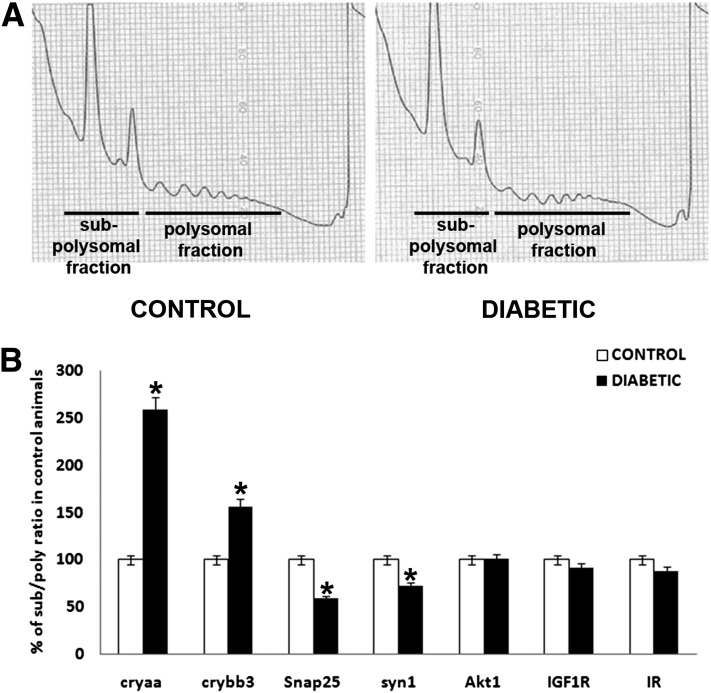
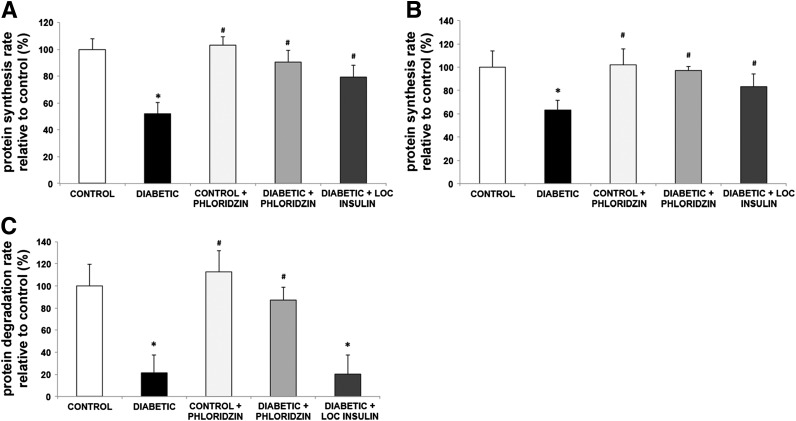
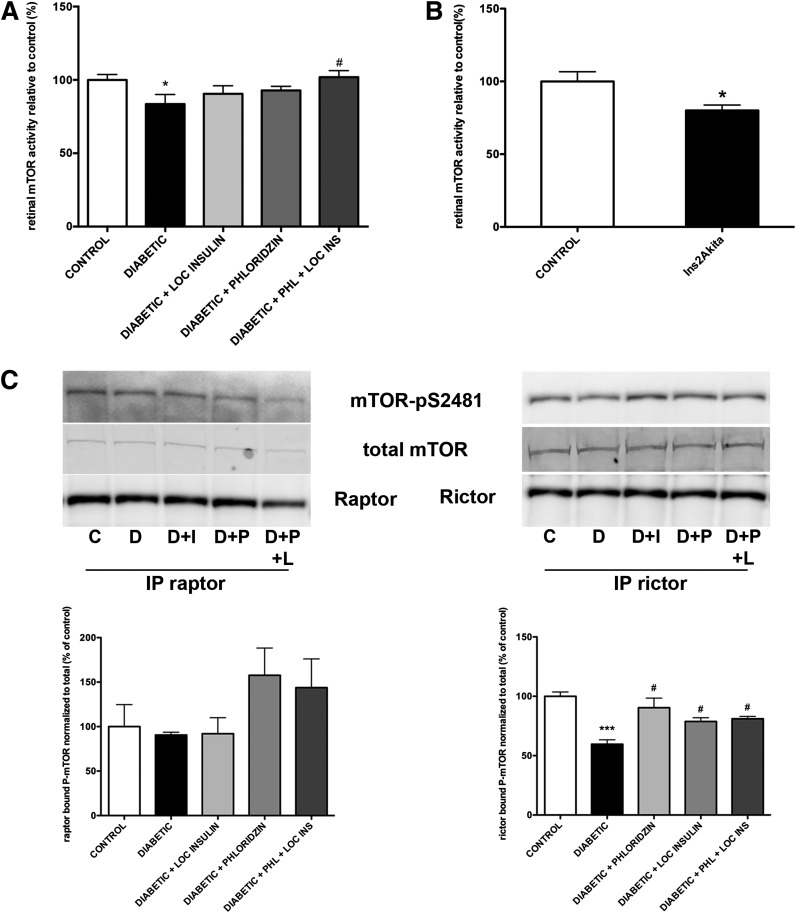
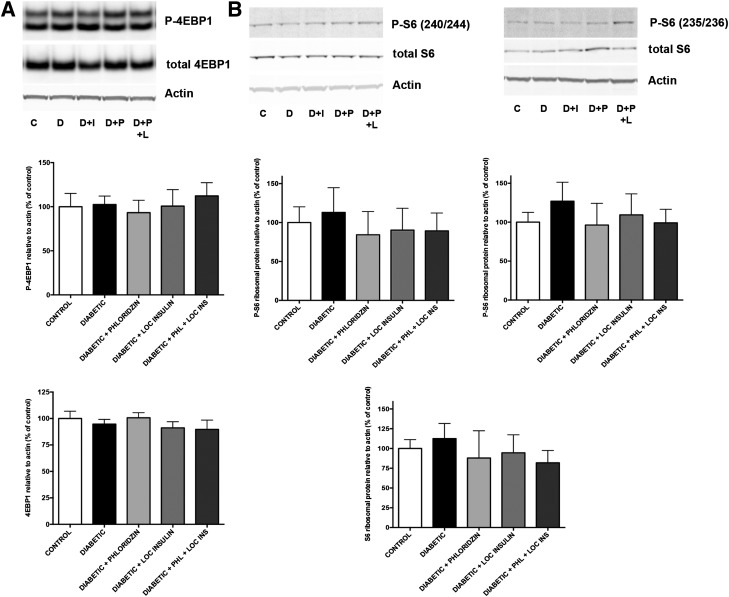
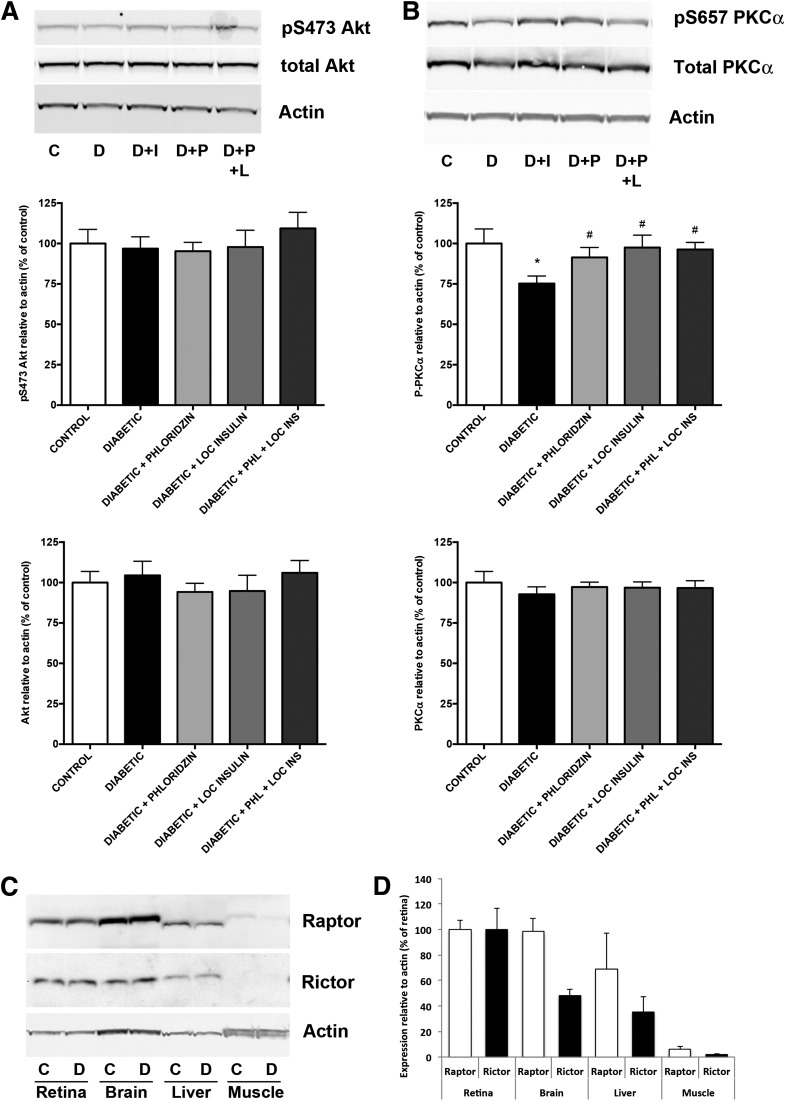
Poorly controlled diabetes has long been known as a catabolic disorder with profound loss of muscle and fat body mass resulting from a simultaneous reduction in protein synthesis and enhanced protein degradation. By contrast, retinal structure is largely maintained during diabetes despite reduced Akt activity and increased rate of cell death. Therefore, we hypothesized that retinal protein turnover is regulated differently than in other insulin-sensitive tissues, such as skeletal muscle. Ins2(Akita) diabetic mice and streptozotocin-induced diabetic rats exhibited marked reductions in retinal protein synthesis matched by a concomitant reduction in retinal protein degradation associated with preserved retinal mass and protein content. The reduction in protein synthesis depended on both hyperglycemia and insulin deficiency, but protein degradation was only reversed by normalization of hyperglycemia. The reduction in protein synthesis was associated with diminished protein translation efficiency but, surprisingly, not with reduced activity of the mTORC1/S6K1/4E-BP1 pathway. Instead, diabetes induced a specific reduction of mTORC2 complex activity. These findings reveal distinctive responses of diabetes-induced retinal protein turnover compared with muscle and liver that may provide a new means to ameliorate diabetic retinopathy.
© 2014 by the American Diabetes Association. Readers may use this article as long as the work is properly cited, the use is educational and not for profit, and the work is not altered.
Figures
References
-
- Antonetti DA, Klein R, Gardner TW. Diabetic retinopathy. N Engl J Med 2012;366:1227–1239 - PubMed
-
- Antonetti DA, Barber AJ, Bronson SK, et al. JDRF Diabetic Retinopathy Center Group Diabetic retinopathy: seeing beyond glucose-induced microvascular disease. Diabetes 2006;55:2401–2411 - PubMed
-
- Ahmed J, Braun RD, Dunn R, Jr, Linsenmeier RA. Oxygen distribution in the macaque retina. Invest Ophthalmol Vis Sci 1993;34:516–521 - PubMed
-
- Wangsa-Wirawan ND, Linsenmeier RA. Retinal oxygen: fundamental and clinical aspects. Arch Ophthalmol 2003;121:547–557 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01 DK015658/DK/NIDDK NIH HHS/United States
- EY-020895/EY/NEI NIH HHS/United States
- P30 DK092926/DK/NIDDK NIH HHS/United States
- R01 EY020582/EY/NEI NIH HHS/United States
- DK-13499/DK/NIDDK NIH HHS/United States
- R01 DK013499/DK/NIDDK NIH HHS/United States
- EY-20582/EY/NEI NIH HHS/United States
- P30 DK020572/DK/NIDDK NIH HHS/United States
- P30 EY007003/EY/NEI NIH HHS/United States
- F31 EY007003/EY/NEI NIH HHS/United States
- DK-097153/DK/NIDDK NIH HHS/United States
- DK-15658/DK/NIDDK NIH HHS/United States
- DP3 DK094292/DK/NIDDK NIH HHS/United States
- R01 EY020895/EY/NEI NIH HHS/United States
- EY-007003/EY/NEI NIH HHS/United States
- DK-094292/DK/NIDDK NIH HHS/United States
- U24 DK097153/DK/NIDDK NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
