

Reprogramming of the chick retinal pigmented epithelium after retinal injury
- PMID: 24742279
- PMCID: PMC4026860
- DOI: 10.1186/1741-7007-12-28
Reprogramming of the chick retinal pigmented epithelium after retinal injury
Abstract
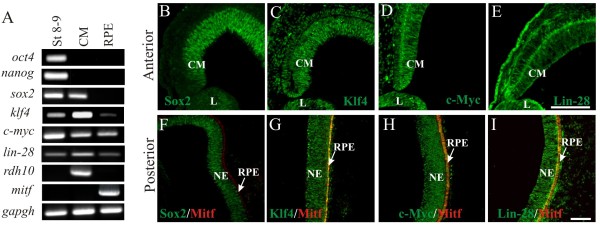
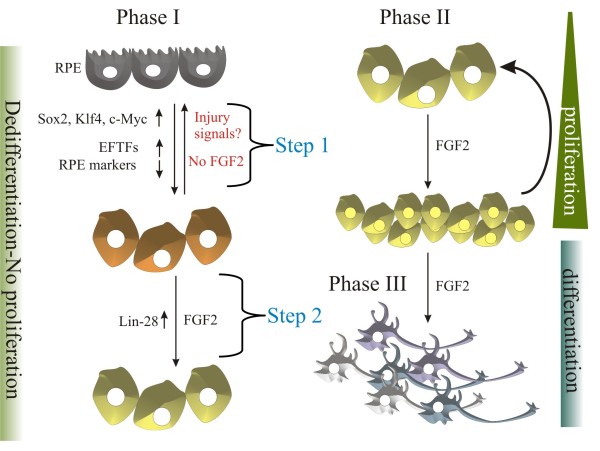
Background: One of the promises in regenerative medicine is to regenerate or replace damaged tissues. The embryonic chick can regenerate its retina by transdifferentiation of the retinal pigmented epithelium (RPE) and by activation of stem/progenitor cells present in the ciliary margin. These two ways of regeneration occur concomitantly when an external source of fibroblast growth factor 2 (FGF2) is present after injury (retinectomy). During the process of transdifferentiation, the RPE loses its pigmentation and is reprogrammed to become neuroepithelium, which differentiates to reconstitute the different cell types of the neural retina. Somatic mammalian cells can be reprogrammed to become induced pluripotent stem cells by ectopic expression of pluripotency-inducing factors such as Oct4, Sox2, Klf4, c-Myc and in some cases Nanog and Lin-28. However, there is limited information concerning the expression of these factors during natural regenerative processes. Organisms that are able to regenerate their organs could share similar mechanisms and factors with the reprogramming process of somatic cells. Herein, we investigate the expression of pluripotency-inducing factors in the RPE after retinectomy (injury) and during transdifferentiation in the presence of FGF2.
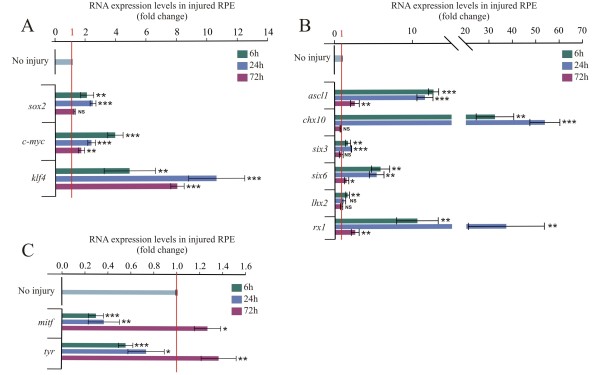
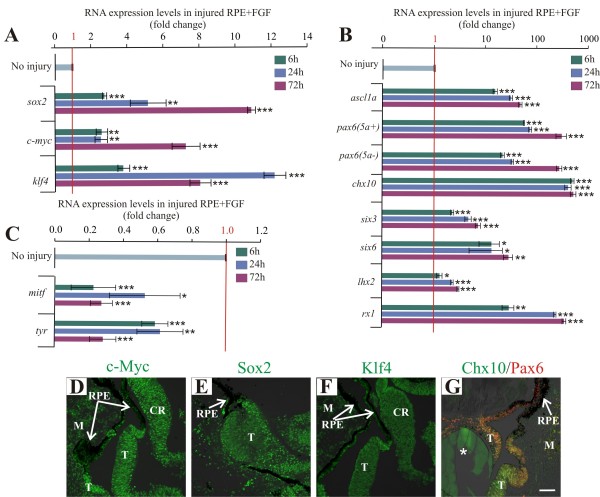
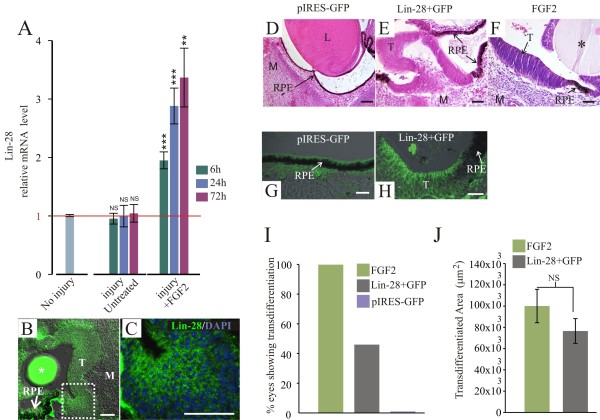
Results: We present evidence that upon injury, the quiescent (p27(Kip1)+/BrdU-) RPE cells transiently dedifferentiate and express sox2, c-myc and klf4 along with eye field transcriptional factors and display a differential up-regulation of alternative splice variants of pax6. However, this transient process of dedifferentiation is not sustained unless FGF2 is present. We have identified lin-28 as a downstream target of FGF2 during the process of retina regeneration. Moreover, we show that overexpression of lin-28 after retinectomy was sufficient to induce transdifferentiation of the RPE in the absence of FGF2.
Conclusion: These findings delineate in detail the molecular changes that take place in the RPE during the process of transdifferentiation in the embryonic chick, and specifically identify Lin-28 as an important factor in this process. We propose a novel model in which injury signals initiate RPE dedifferentiation, while FGF2 up-regulates Lin-28, allowing for RPE transdifferentiation to proceed.
Figures

References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
