

Representing sex in the brain, one module at a time
- PMID: 24742456
- PMCID: PMC4130170
- DOI: 10.1016/j.neuron.2014.03.029
Representing sex in the brain, one module at a time
Abstract
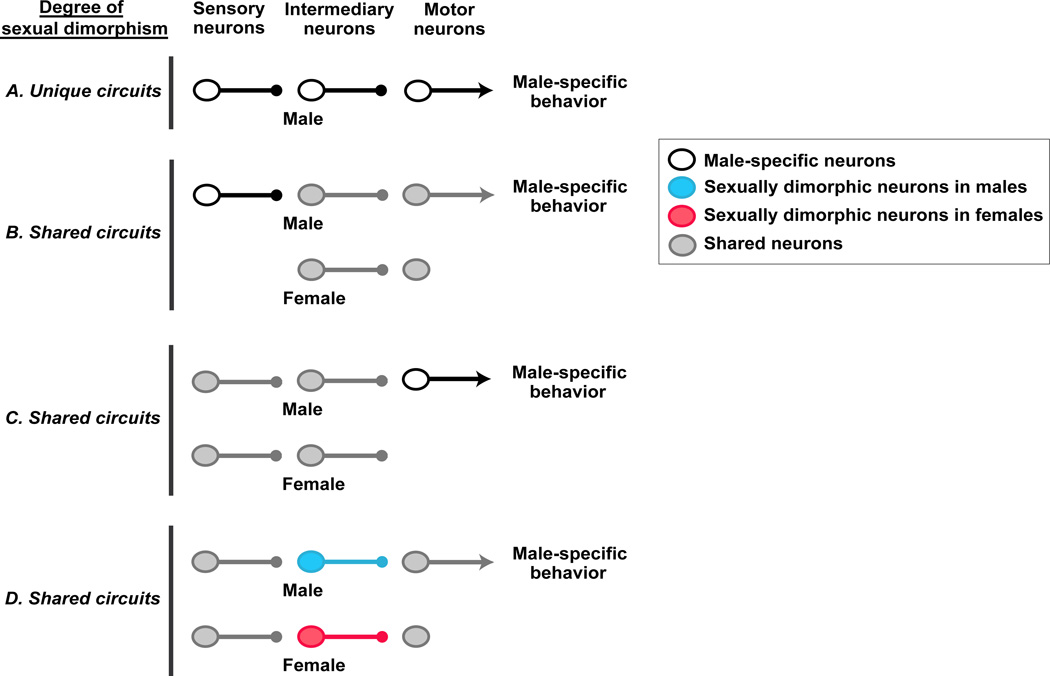
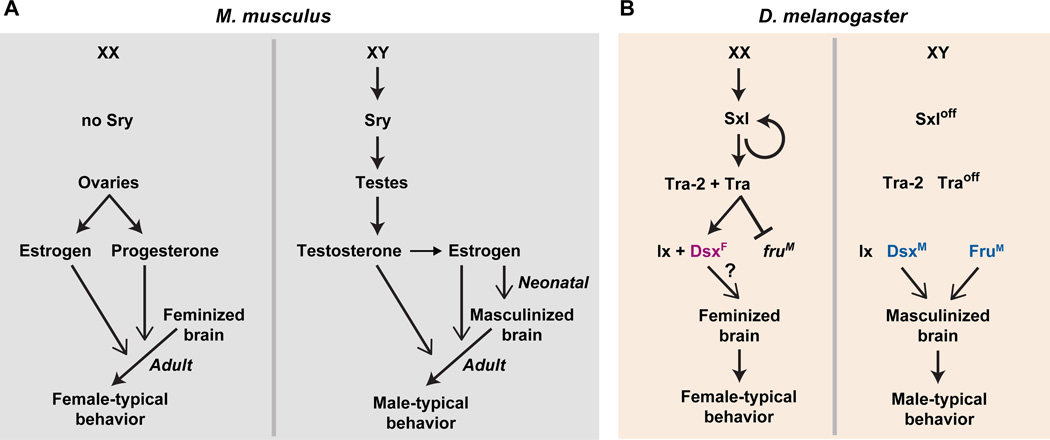
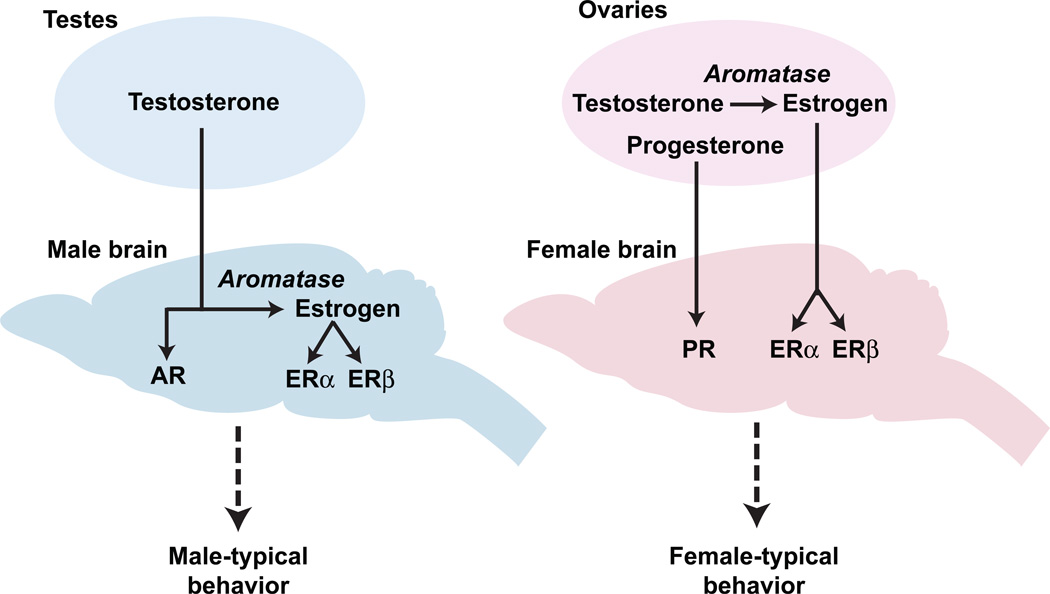
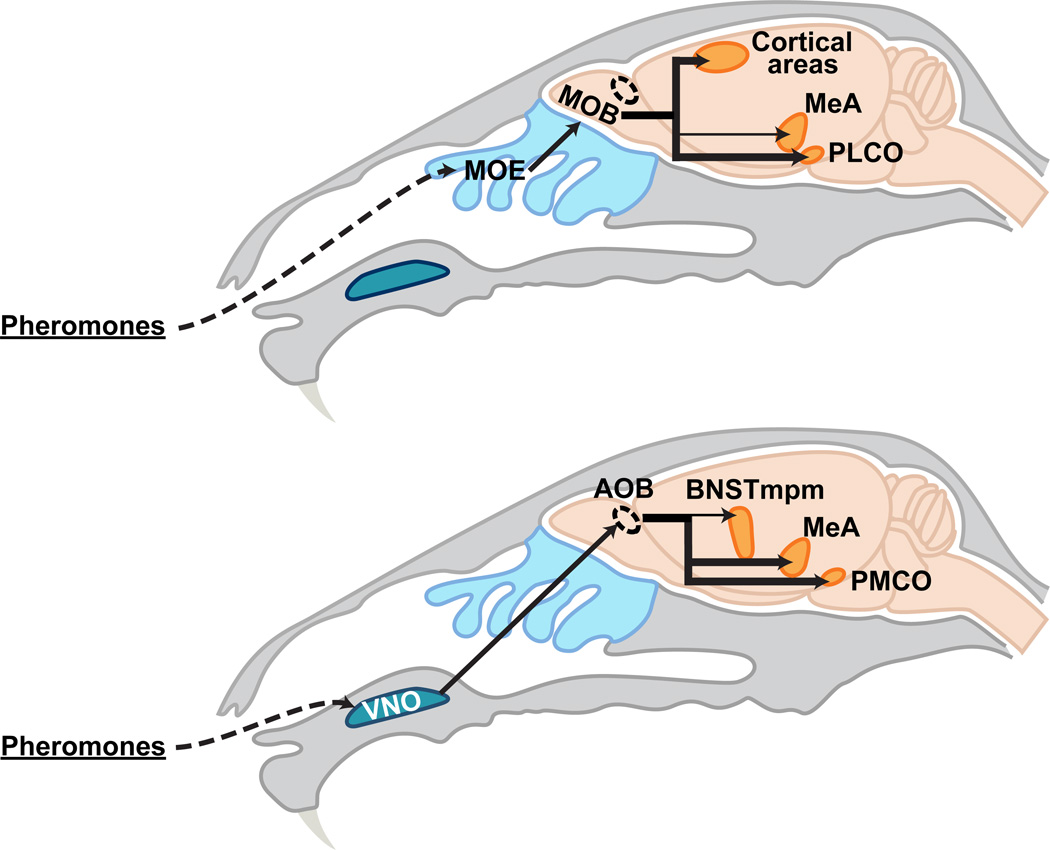
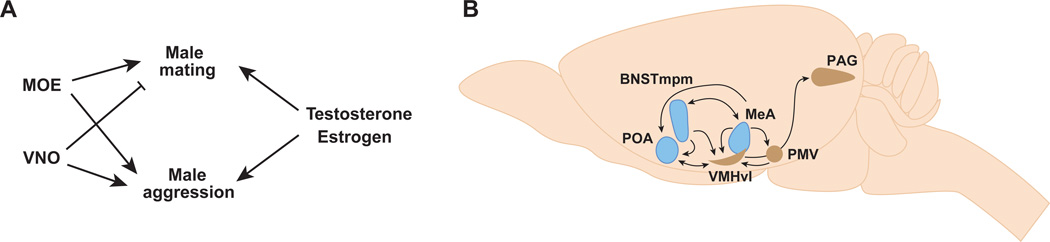
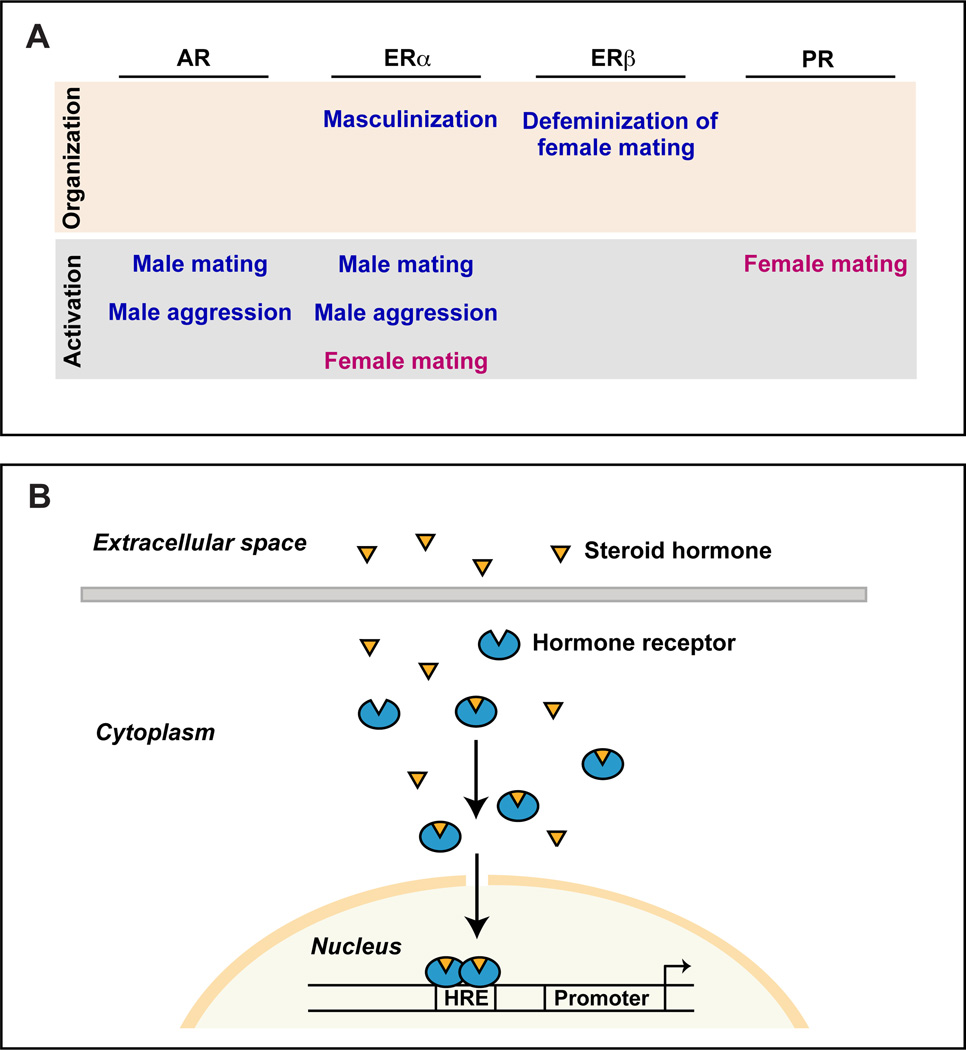
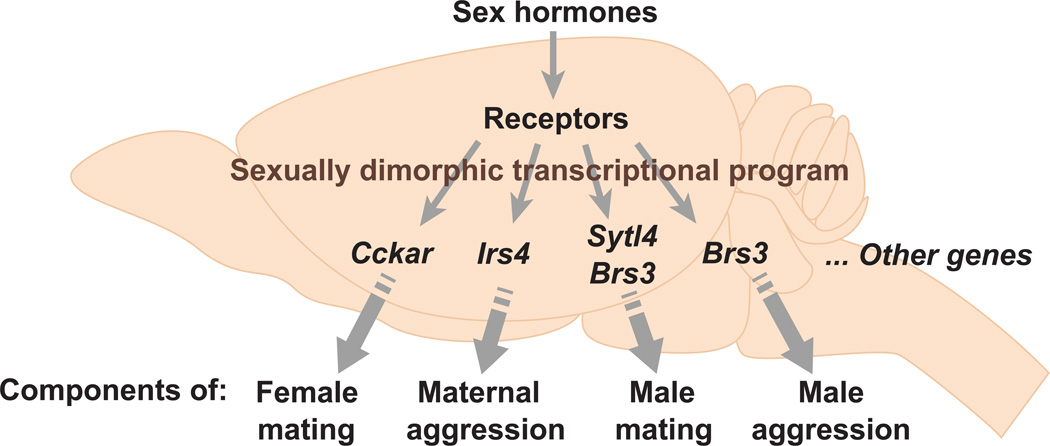
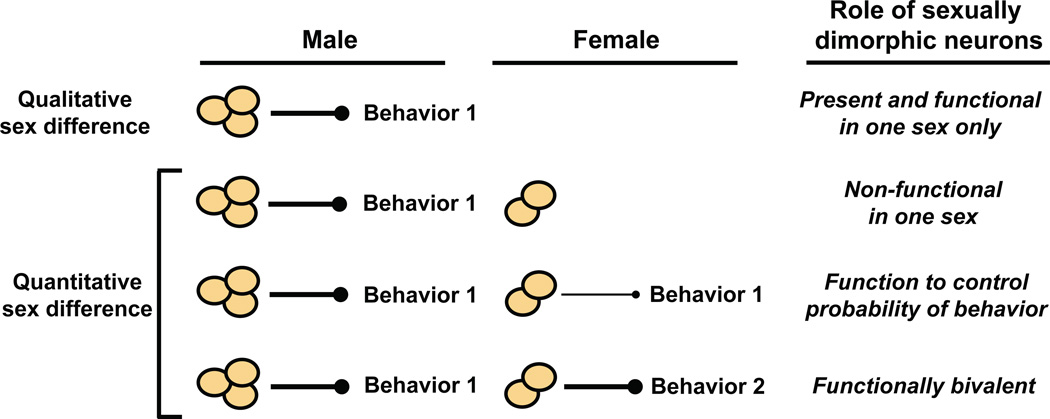
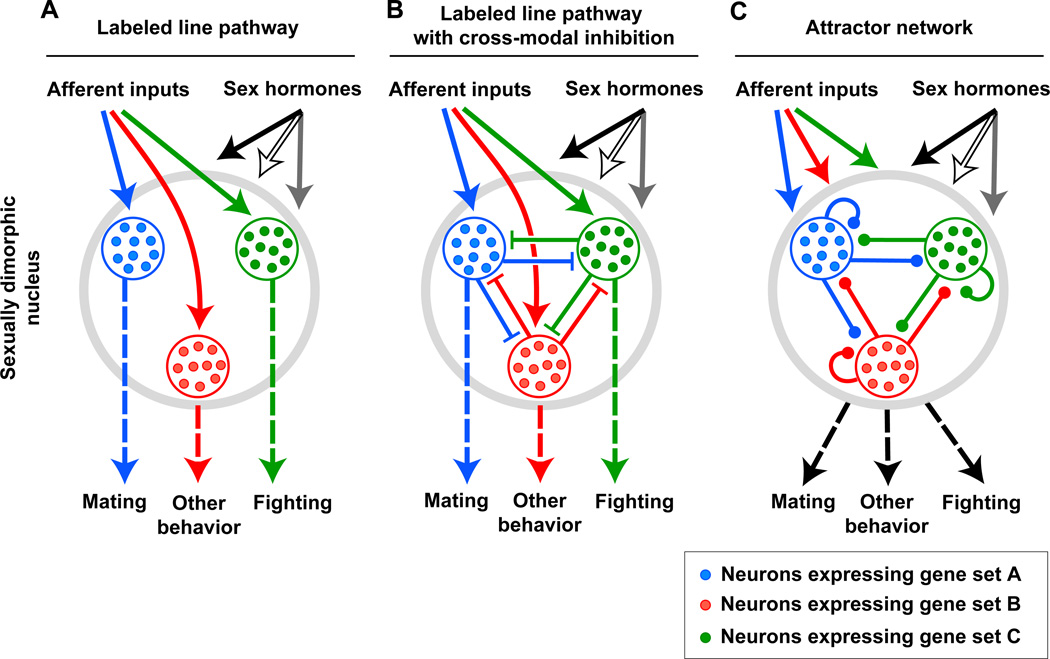
Sexually dimorphic behaviors, qualitative or quantitative differences in behaviors between the sexes, result from the activity of a sexually differentiated nervous system. Sensory cues and sex hormones control the entire repertoire of sexually dimorphic behaviors, including those commonly thought to be charged with emotion such as courtship and aggression. Such overarching control mechanisms regulate distinct genes and neurons that in turn specify the display of these behaviors in a modular manner. How such modular control is transformed into cohesive internal states that correspond to sexually dimorphic behavior is poorly understood. We summarize current understanding of the neural circuit control of sexually dimorphic behaviors from several perspectives, including how neural circuits in general, and sexually dimorphic neurons in particular, can generate sexually dimorphic behaviors, and how molecular mechanisms and evolutionary constraints shape these behaviors. We propose that emergent themes such as the modular genetic and neural control of dimorphic behavior are broadly applicable to the neural control of other behaviors.
Copyright © 2014 Elsevier Inc. All rights reserved.
Figures
References
-
- Allen E, Francis BF, Robertson LL, Colgate CE, Johnston CG, Doisy EA, Kountz WB, Gibson HV. The hormone of the ovarian follicle; its localization and action in test animals, and additional points bearing upon the internal secretion of the ovary. Am. J. Anat. 1924;34:133–181.
-
- Amateau SK, Alt JJ, Stamps CL, McCarthy MM. Brain estradiol content in newborn rats: sex differences, regional heterogeneity, and possible de novo synthesis by the female telencephalon. Endocrinology. 2004;145:2906–2917. - PubMed
-
- Antliff HR, Young WC. Behavioral and tissue responses of male guinea pigs to estrogens and the problem of hormone specificity. Endocrinology. 1956;59:74–82. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
