

Α-tubulin K40 acetylation is required for contact inhibition of proliferation and cell-substrate adhesion
- PMID: 24743598
- PMCID: PMC4055265
- DOI: 10.1091/mbc.E13-10-0609
Α-tubulin K40 acetylation is required for contact inhibition of proliferation and cell-substrate adhesion
Abstract
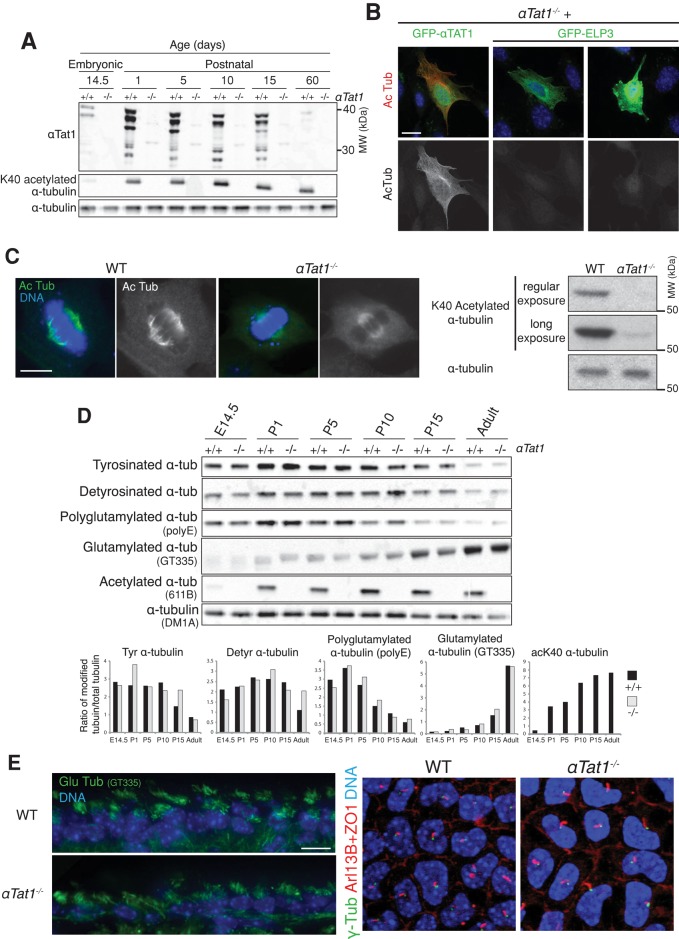
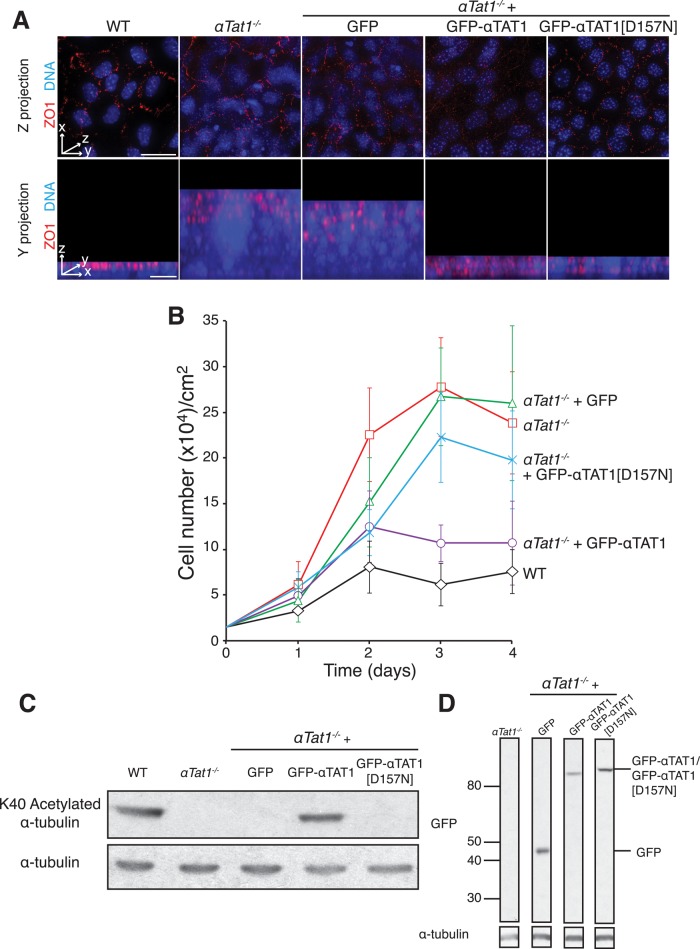
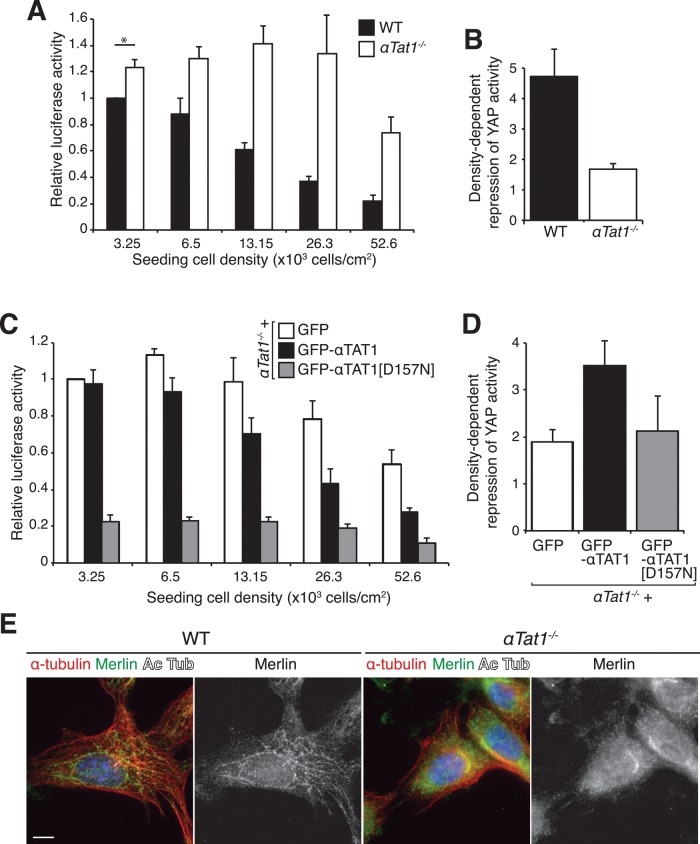
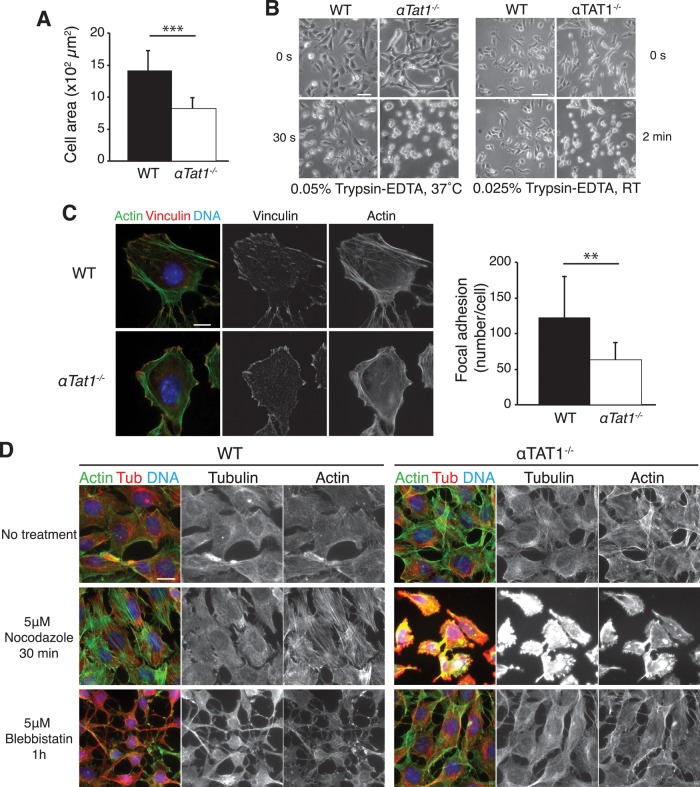
Acetylation of α-tubulin on lysine 40 marks long-lived microtubules in structures such as axons and cilia, and yet the physiological role of α-tubulin K40 acetylation is elusive. Although genetic ablation of the α-tubulin K40 acetyltransferase αTat1 in mice did not lead to detectable phenotypes in the developing animals, contact inhibition of proliferation and cell-substrate adhesion were significantly compromised in cultured αTat1(-/-) fibroblasts. First, αTat1(-/-) fibroblasts kept proliferating beyond the confluent monolayer stage. Congruently, αTat1(-/-) cells failed to activate Hippo signaling in response to increased cell density, and the microtubule association of the Hippo regulator Merlin was disrupted. Second, αTat1(-/-) cells contained very few focal adhesions, and their ability to adhere to growth surfaces was greatly impaired. Whereas the catalytic activity of αTAT1 was dispensable for monolayer formation, it was necessary for cell adhesion and restrained cell proliferation and activation of the Hippo pathway at elevated cell density. Because α-tubulin K40 acetylation is largely eliminated by deletion of αTAT1, we propose that acetylated microtubules regulate contact inhibition of proliferation through the Hippo pathway.
© 2014 Aguilar et al. This article is distributed by The American Society for Cell Biology under license from the author(s). Two months after publication it is available to the public under an Attribution–Noncommercial–Share Alike 3.0 Unported Creative Commons License (http://creativecommons.org/licenses/by-nc-sa/3.0).
Figures
References
-
- Abercrombie M. Contact-dependent behavior of normal cells and the possible significance of surface changes in virus-induced transformation. Cold Spring Harb Symp Quant Biol. 1962;27:427–431. - PubMed
-
- Abercrombie M. Contact inhibition and malignancy. Nature. 1979;281:259–262. - PubMed
-
- Aragona M, Panciera T, Manfrin A, Giulitti S, Michielin F, Elvassore N, Dupont S, Piccolo S. A mechanical checkpoint controls multicellular growth through YAP/TAZ regulation by actin-processing factors. Cell. 2013;154:1047–1059. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
