

Exposure-dependent control of malaria-induced inflammation in children
- PMID: 24743880
- PMCID: PMC3990727
- DOI: 10.1371/journal.ppat.1004079
Exposure-dependent control of malaria-induced inflammation in children
Abstract
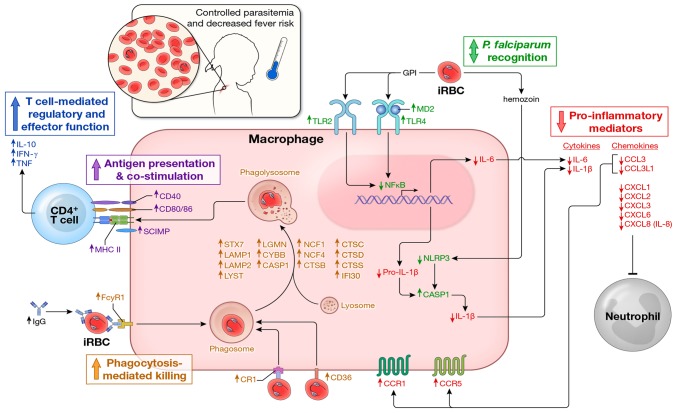
In malaria-naïve individuals, Plasmodium falciparum infection results in high levels of parasite-infected red blood cells (iRBCs) that trigger systemic inflammation and fever. Conversely, individuals in endemic areas who are repeatedly infected are often asymptomatic and have low levels of iRBCs, even young children. We hypothesized that febrile malaria alters the immune system such that P. falciparum re-exposure results in reduced production of pro-inflammatory cytokines/chemokines and enhanced anti-parasite effector responses compared to responses induced before malaria. To test this hypothesis we used a systems biology approach to analyze PBMCs sampled from healthy children before the six-month malaria season and the same children seven days after treatment of their first febrile malaria episode of the ensuing season. PBMCs were stimulated with iRBC in vitro and various immune parameters were measured. Before the malaria season, children's immune cells responded to iRBCs by producing pro-inflammatory mediators such as IL-1β, IL-6 and IL-8. Following malaria there was a marked shift in the response to iRBCs with the same children's immune cells producing lower levels of pro-inflammatory cytokines and higher levels of anti-inflammatory cytokines (IL-10, TGF-β). In addition, molecules involved in phagocytosis and activation of adaptive immunity were upregulated after malaria as compared to before. This shift was accompanied by an increase in P. falciparum-specific CD4+Foxp3- T cells that co-produce IL-10, IFN-γ and TNF; however, after the subsequent six-month dry season, a period of markedly reduced malaria transmission, P. falciparum-inducible IL-10 production remained partially upregulated only in children with persistent asymptomatic infections. These findings suggest that in the face of P. falciparum re-exposure, children acquire exposure-dependent P. falciparum-specific immunoregulatory responses that dampen pathogenic inflammation while enhancing anti-parasite effector mechanisms. These data provide mechanistic insight into the observation that P. falciparum-infected children in endemic areas are often afebrile and tend to control parasite replication.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures



References
-
- Collins WE, Jeffery GM (1999) A retrospective examination of sporozoite- and trophozoite-induced infections with Plasmodium falciparum: development of parasitologic and clinical immunity during primary infection. Am J Trop Med Hyg 61: 4–19. - PubMed
-
- Day NP, Hien TT, Schollaardt T, Loc PP, Chuong LV, et al. (1999) The prognostic and pathophysiologic role of pro- and antiinflammatory cytokines in severe malaria. J Infect Dis 180: 1288–1297. - PubMed
-
- Lyke KE, Burges R, Cissoko Y, Sangare L, Dao M, et al. (2004) Serum levels of the proinflammatory cytokines interleukin-1 beta (IL-1beta), IL-6, IL-8, IL-10, tumor necrosis factor alpha, and IL-12(p70) in Malian children with severe Plasmodium falciparum malaria and matched uncomplicated malaria or healthy controls. Infect Immun 72: 5630–5637. - PMC - PubMed
-
- Walther M, Woodruff J, Edele F, Jeffries D, Tongren JE, et al. (2006) Innate immune responses to human malaria: heterogeneous cytokine responses to blood-stage Plasmodium falciparum correlate with parasitological and clinical outcomes. J Immunol 177: 5736–5745. - PubMed
-
- Urban BC, Ing R, Stevenson MM (2005) Early interactions between blood-stage plasmodium parasites and the immune system. Curr Top Microbiol Immunol 297: 25–70. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
