

Functional analysis of the dioxin response elements (DREs) of the murine CYP1A1 gene promoter: beyond the core DRE sequence
- PMID: 24743890
- PMCID: PMC4013641
- DOI: 10.3390/ijms15046475
Functional analysis of the dioxin response elements (DREs) of the murine CYP1A1 gene promoter: beyond the core DRE sequence
Abstract
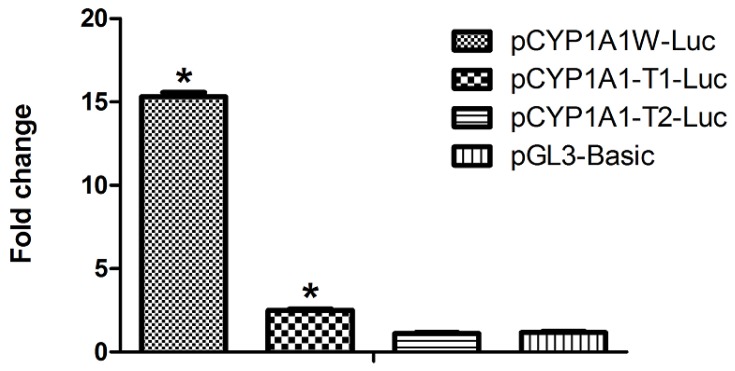
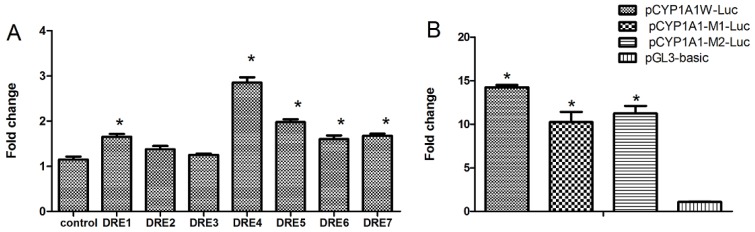
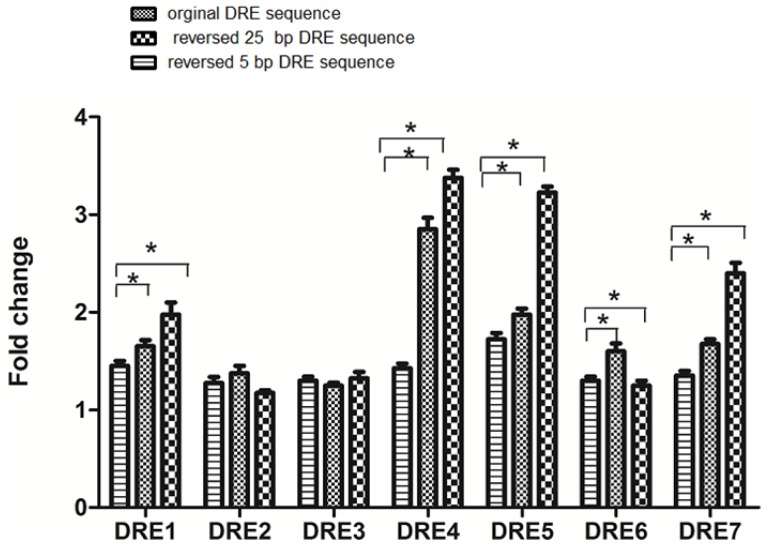
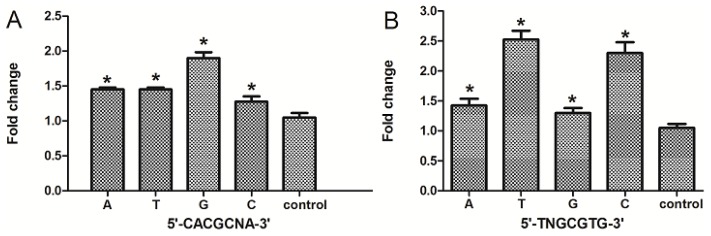
The aryl hydrocarbon receptor (AhR) is a ligand-dependent transcription factor that mediates the biological and toxicological effects of halogenated aromatic hydrocarbons, such as 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD). When activated by dioxin, the cytosolic AhR protein complex translocates into the nucleus and dimerizes with the ARNT (Ah receptor nuclear translocator) protein. The heteromeric ligand:AhR/Arnt complex then recognizes and binds to its specific DNA recognition site, the dioxin response element (DRE). DREs are located upstream of cytochrome P4501A1 (CYP1A1) and other AhR-responsive genes, and binding of the AhR complex stimulates their transcription. Although CYP1A1 expression has been used as the model system to define the biochemical and molecular mechanism of AhR action, there is still limited knowledge about the roles of each of the seven DREs located in the CYP1A1 promoter. These seven DREs are conserved in mouse, human and rat. Deletion analysis showed that a single DRE at -488 was enough to activate the transcription. Truncation analysis demonstrated that the DRE at site -981 has the highest transcriptional efficiency in response to TCDD. This result was verified by mutation analysis, suggesting that the conserved DRE at site -981 could represent a significant and universal AhR regulatory element for CYP1A1. The reversed substituted intolerant core sequence (5'-GCGTG-3' or 5'-CACGC-3') of seven DREs reduced the transcriptional efficiency, which illustrated that the adjacent sequences of DRE played a vital role in activating transcription. The core DRE sequence (5'-TNGCGTG-3') tends to show a higher transcriptional level than that of the core DRE sequence (5'-CACGCNA-3') triggered by TCDD. Furthermore, in the core DRE (5'-TNGCGTG-3') sequence, when "N" is thymine or cytosine (T or C), the transcription efficiency was stronger compared with that of the other nucleotides. The effects of DRE orientation, DRE adjacent sequences and the nucleotide "N" in the core DRE (5'-TNGCGTG-3') sequence on the AhR-regulated CYP1A1 transcription in response to TCDD were studied systematically, and our study laid a good foundation for further investigation into the AhR-dependent transcriptional regulation triggered by dioxin and dioxin-like compounds.
Figures

References
-
- Hankinson O. The aryl hydrocarbon receptor complex. Annu. Rev. Pharmacol. Toxicol. 1995;35:307–340. - PubMed
-
- Safe S. Polychlorinated biphenyls (PCBs), dibenzo-p-dioxins (PCDDs), dibenzofurans (PCDFs), and related compounds: Environmental and mechanistic considerations which support the development of toxic equivalency factors (TEFs) Crit. Rev. Toxicol. 1990;21:51–88. - PubMed
-
- Whitlock J.P., Jr. Induction of cytochrome P4501A1. Annu. Rev. Pharmacol. Toxicol. 1999;39:103–125. - PubMed
-
- Senft A.P., Daltion T.P., Nebert D.W., Genter M.B., Hutchinson R.J., Schertzer H.G. Dioxin increases reactive oxygen production in mouse liver mitochondria. Toxicol. Appl. Pharmacol. 2002;178:15–21. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
