

Circularly permuted tRNA genes: their expression and implications for their physiological relevance and development
- PMID: 24744771
- PMCID: PMC3978253
- DOI: 10.3389/fgene.2014.00063
Circularly permuted tRNA genes: their expression and implications for their physiological relevance and development
Abstract
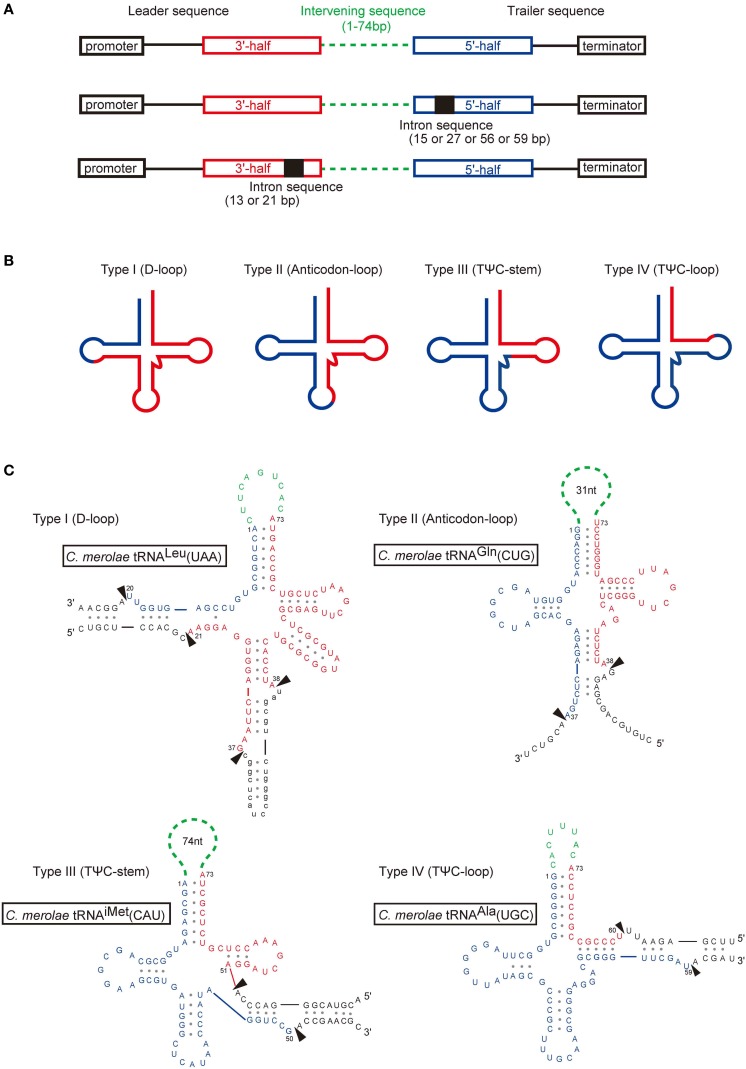
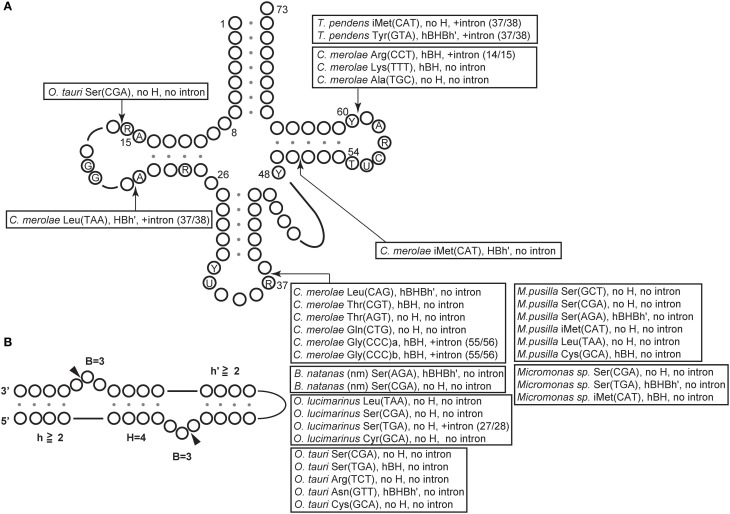
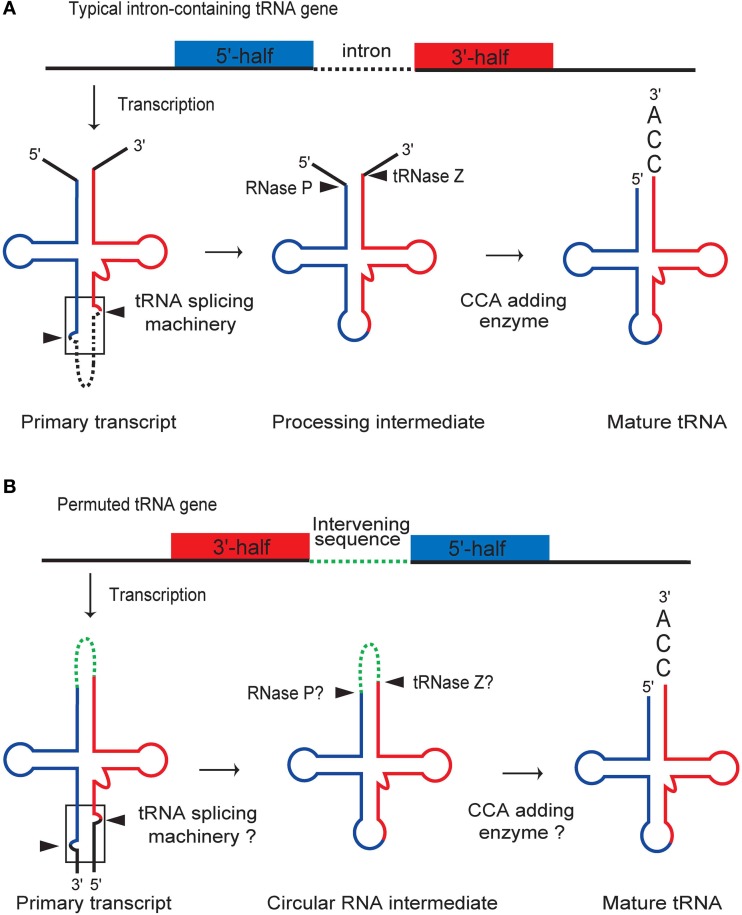
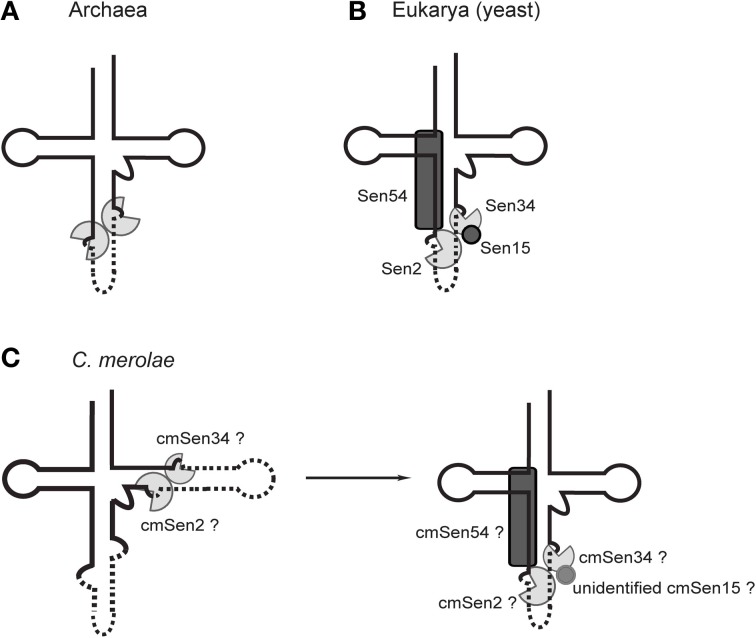
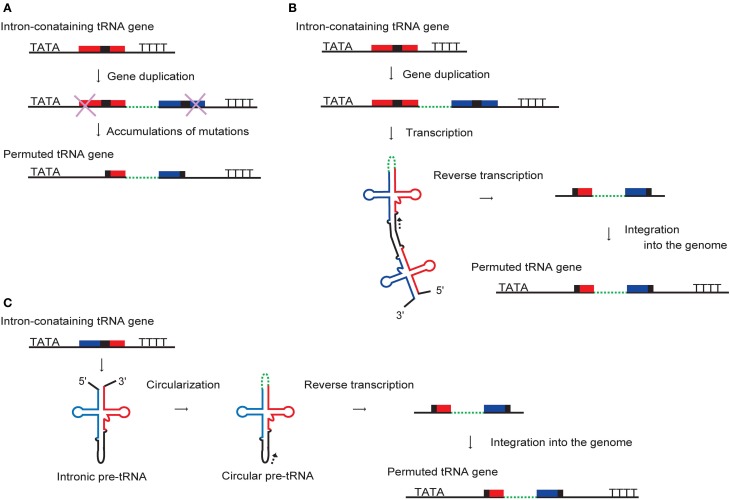
A number of genome analyses and searches using programs that focus on the RNA-specific bulge-helix-bulge (BHB) motif have uncovered a wide variety of disrupted tRNA genes. The results of these analyses have shown that genetic information encoding functional RNAs is described in the genome cryptically and is retrieved using various strategies. One such strategy is represented by circularly permuted tRNA genes, in which the sequences encoding the 5'-half and 3'-half of the specific tRNA are separated and inverted on the genome. Biochemical analyses have defined a processing pathway in which the termini of tRNA precursors (pre-tRNAs) are ligated to form a characteristic circular RNA intermediate, which is then cleaved at the acceptor-stem to generate the typical cloverleaf structure with functional termini. The sequences adjacent to the processing site located between the 3'-half and the 5'-half of pre-tRNAs potentially form a BHB motif, which is the dominant recognition site for the tRNA-intron splicing endonuclease, suggesting that circularization of pre-tRNAs depends on the splicing machinery. Some permuted tRNAs contain a BHB-mediated intron in their 5'- or 3'-half, meaning that removal of an intron, as well as swapping of the 5'- and 3'-halves, are required during maturation of their pre-tRNAs. To date, 34 permuted tRNA genes have been identified from six species of unicellular algae and one archaeon. Although their physiological significance and mechanism of development remain unclear, the splicing system of BHB motifs seems to have played a key role in the formation of permuted tRNA genes. In this review, current knowledge of circularly permuted tRNA genes is presented and some unanswered questions regarding these species are discussed.
Keywords: BHB motif; circular gene permutation; intron; tRNA gene; tRNA-splicing endonuclease.
Figures
Similar articles
-
Permuted tRNA genes in the nuclear and nucleomorph genomes of photosynthetic eukaryotes.Mol Biol Evol. 2010 May;27(5):1070-6. doi: 10.1093/molbev/msp313. Epub 2009 Dec 18. Mol Biol Evol. 2010. PMID: 20022888
-
Identification of highly-disrupted tRNA genes in nuclear genome of the red alga, Cyanidioschyzon merolae 10D.Sci Rep. 2013;3:2321. doi: 10.1038/srep02321. Sci Rep. 2013. PMID: 23900518 Free PMC article.
-
Avatar pre-tRNAs help elucidate the properties of tRNA-splicing endonucleases that produce tRNA from permuted genes.Proc Natl Acad Sci U S A. 2012 Dec 26;109(52):21325-9. doi: 10.1073/pnas.1219336110. Epub 2012 Dec 11. Proc Natl Acad Sci U S A. 2012. PMID: 23236183 Free PMC article.
-
tRNA gene diversity in the three domains of life.Front Genet. 2014 May 26;5:142. doi: 10.3389/fgene.2014.00142. eCollection 2014. Front Genet. 2014. PMID: 24904642 Free PMC article. Review.
-
Intron-dependent enzymatic formation of modified nucleosides in eukaryotic tRNAs: a review.Biochimie. 1997 May;79(5):293-302. doi: 10.1016/s0300-9084(97)83517-1. Biochimie. 1997. PMID: 9258438 Review.
Cited by
-
Welcome to the new tRNA world!Front Genet. 2014 Sep 23;5:336. doi: 10.3389/fgene.2014.00336. eCollection 2014. Front Genet. 2014. PMID: 25295055 Free PMC article. No abstract available.
-
Circular RNAs: A novel type of non-coding RNA and their potential implications in antiviral immunity.Int J Biol Sci. 2017 Nov 2;13(12):1497-1506. doi: 10.7150/ijbs.22531. eCollection 2017. Int J Biol Sci. 2017. PMID: 29230098 Free PMC article. Review.
-
Discovery of natural non-circular permutations in non-coding RNAs.Nucleic Acids Res. 2023 Apr 11;51(6):2850-2861. doi: 10.1093/nar/gkad137. Nucleic Acids Res. 2023. PMID: 36912096 Free PMC article.
-
The 23S Ribosomal RNA From Pyrococcus furiosus Is Circularly Permuted.Front Microbiol. 2020 Dec 10;11:582022. doi: 10.3389/fmicb.2020.582022. eCollection 2020. Front Microbiol. 2020. PMID: 33362734 Free PMC article.
-
Circular RNAs: diversity of form and function.RNA. 2014 Dec;20(12):1829-42. doi: 10.1261/rna.047126.114. RNA. 2014. PMID: 25404635 Free PMC article. Review.
References
-
- Altman S., Kirsebom L., Talbot S. (1995). Recent studies of RNase P, in tRNA, eds Söll D., RajBhandary U. L. (Washington, DC: ASM Press; ), 67–78
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources