

Photoconversion changes bilin chromophore conjugation and protein secondary structure in the violet/orange cyanobacteriochrome NpF2164g3' [corrected]
- PMID: 24745038
- PMCID: PMC4062569
- DOI: 10.1039/c3pp50442e
Photoconversion changes bilin chromophore conjugation and protein secondary structure in the violet/orange cyanobacteriochrome NpF2164g3' [corrected]
Erratum in
- Photochem Photobiol Sci. 2014 Sep;13(9):1360
Abstract
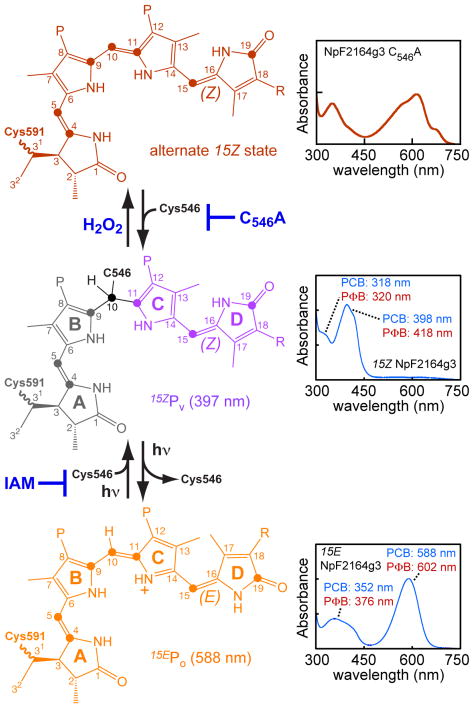
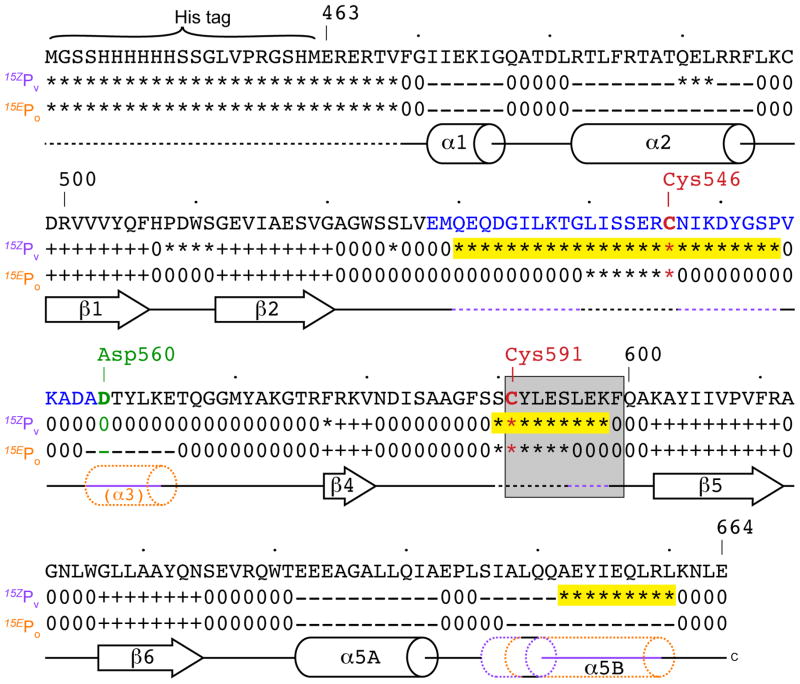
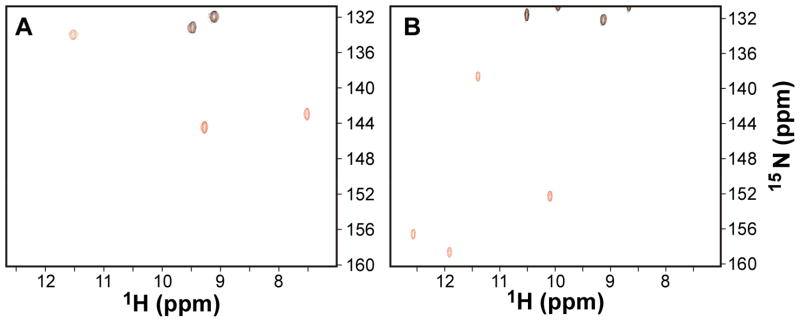
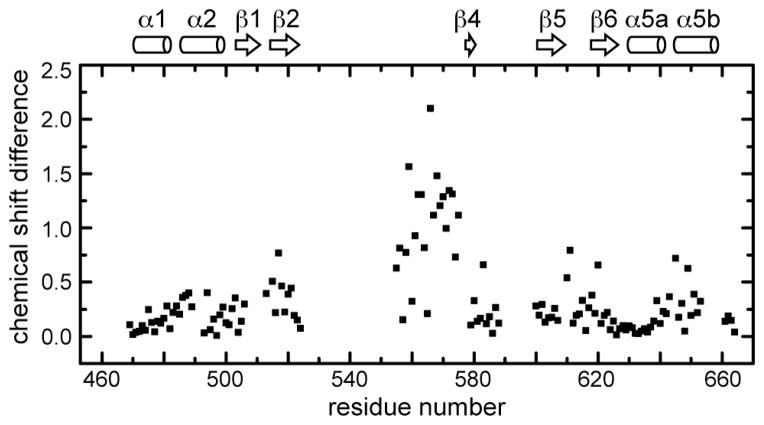
Cyanobacteriochromes (CBCRs) are cyanobacterial photoreceptors distantly related to phytochromes. All CBCRs examined to date utilize a conserved Cys residue to form a covalent thioether linkage to the bilin chromophore. In the insert-Cys CBCR subfamily, a second conserved Cys can covalently link to the bilin C10 methine bridge, allowing detection of near-UV to blue light. The best understood insert-Cys CBCR is the violet/orange CBCR NpF2164g3 from Nostoc punctiforme, which has a stable second linkage in the violet-absorbing dark state. Photoconversion of NpF2164g3 leads to elimination of the second linkage and formation of an orange-absorbing photoproduct. We recently reported NMR chemical shift assignments for the orange-absorbing photoproduct state of NpF2164g3. We here present equivalent information for its violet-absorbing dark state. In both photostates, NpF2164g3 is monomeric in solution and regions containing the two conserved Cys residues essential for photoconversion are structurally disordered. In contrast to blue light receptors such as phototropin, NpF2164g3 is less structurally ordered in the dark state than in the photoproduct. The insert-Cys insertion loop and C-terminal helix exhibit light-dependent structural changes. Moreover, a motif containing an Asp residue also found in other CBCRs and in phytochromes adopts a random-coil structure in the dark state but a stable α-helix structure in the photoproduct. NMR analysis of the chromophore is consistent with a less ordered dark state, with A-ring resonances only resolved in the photoproduct. The C10 atom of the bilin chromophore exhibits a drastic change in chemical shift upon photoconversion, changing from 34.5 ppm (methylene) in the dark state to 115 ppm (methine) in the light-activated state. Our results provide structural insight into the two-Cys photocycle of NpF2164g3 and the structurally diverse mechanisms used for light perception by the larger phytochrome superfamily.
Figures



References
-
- Yeh KC, Wu SH, Murphy JT, Lagarias JC. A cyanobacterial phytochrome two-component light sensory system. Science. 1997;277:1505–1508. - PubMed
-
- Wagner JR, Brunzelle JS, Forest KT, Vierstra RD. A light-sensing knot revealed by the structure of the chromophore binding domain of phytochrome. Nature. 2005;438:325–331. - PubMed
-
- Wagner JR, Zhang J, Brunzelle JS, Vierstra RD, Forest KT. High resolution structure of Deinococcus bacteriophytochrome yields new insights into phytochrome architecture and evolution. J Biol Chem. 2007;282:12298–12309. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources