

Ralstonia solanacearum type III secretion system effector Rip36 induces a hypersensitive response in the nonhost wild eggplant Solanum torvum
- PMID: 24745046
- PMCID: PMC6638889
- DOI: 10.1111/mpp.12079
Ralstonia solanacearum type III secretion system effector Rip36 induces a hypersensitive response in the nonhost wild eggplant Solanum torvum
Abstract
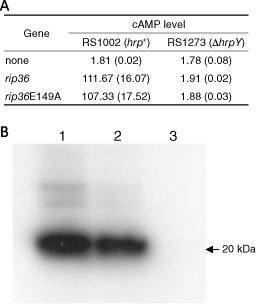
Ralstonia solanacearum is a Gram-negative soil-borne bacterium that causes bacterial wilt disease in more than 200 plant species, including economically important Solanaceae species. In R. solanacearum, the hypersensitive response and pathogenicity (Hrp) type III secretion system is required for both the ability to induce the hypersensitive response (HR) in nonhost plants and pathogenicity in host plants. Recently, 72 effector genes, called rip (Ralstonia protein injected into plant cells), have been identified in R. solanacearum RS1000. RS1002, a spontaneous nalixidic acid-resistant derivative of RS1000, induced strong HR in the nonhost wild eggplant Solanum torvum in an Hrp-dependent manner. An Agrobacterium-mediated transient expression system revealed that Rip36, a putative Zn-dependent protease effector of R. solanacearum, induced HR in S. torvum. A mutation in the putative Zn-binding motif (E149A) completely abolished the ability to induce HR. In agreement with this result, the RS1002-derived Δrip36 and rip36E149A mutants lost the ability to induce HR in S. torvum. An E149A mutation had no effect on the translocation of Rip36 into plant cells. These results indicate that Rip36 is an avirulent factor that induces HR in S. torvum and that a putative Zn-dependent protease motif is essential for this activity.
Figures


References
-
- Alexeyev, M.F. , Shokolenko, I.N. and Croughan, T.P. (1995) New mini‐Tn5 derivatives for insertion mutagenesis and genetic engineering in gram‐negative bacteria. Can. J. Microbiol. 41, 1053–1055. - PubMed
-
- Axtell, M.J. and Staskawicz, B.J. (2003) Initiation of RPS2‐specified disease resistance in Arabidopsis is coupled to the AvrRpt2‐directed elimination of RIN4. Cell, 112, 369–377. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
