

Cyclin B1/Cdk1 coordinates mitochondrial respiration for cell-cycle G2/M progression
- PMID: 24746669
- PMCID: PMC4156313
- DOI: 10.1016/j.devcel.2014.03.012
Cyclin B1/Cdk1 coordinates mitochondrial respiration for cell-cycle G2/M progression
Abstract
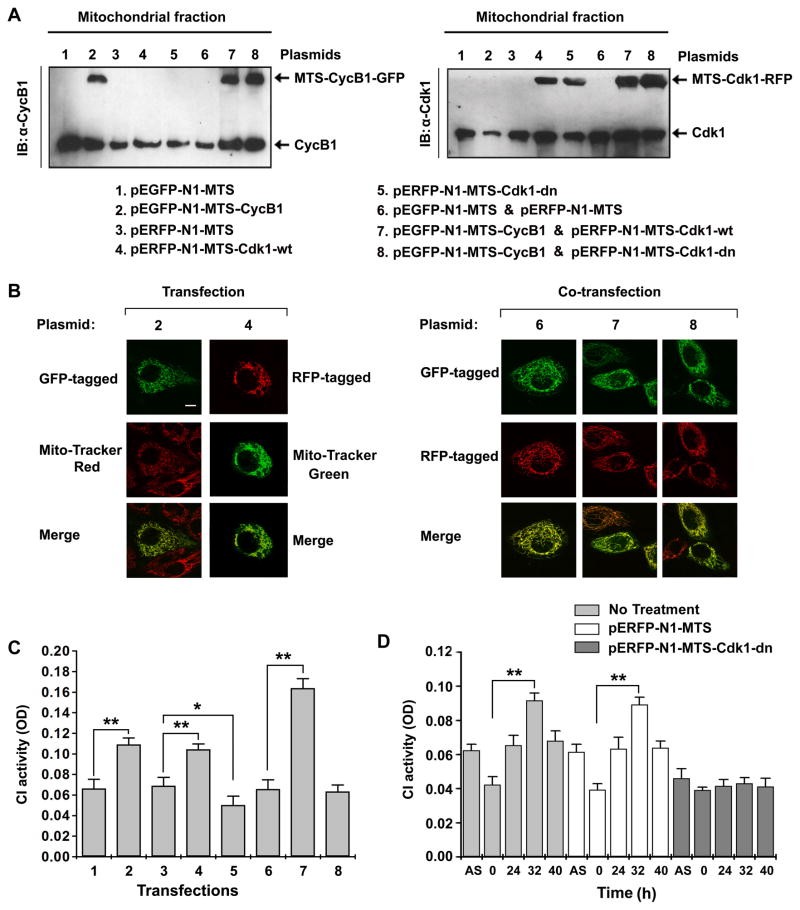
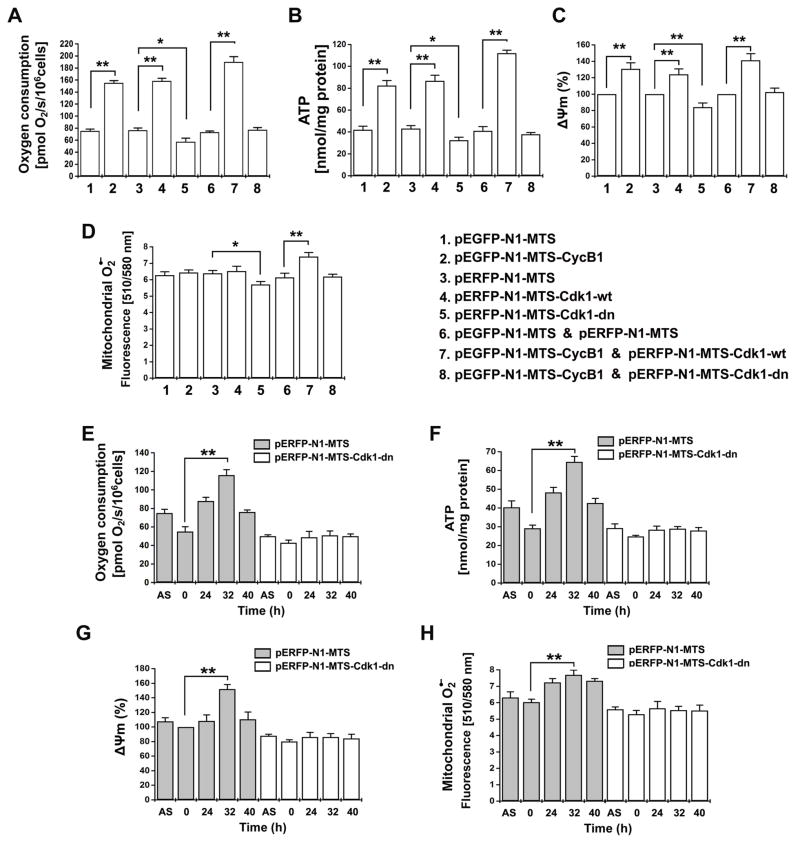
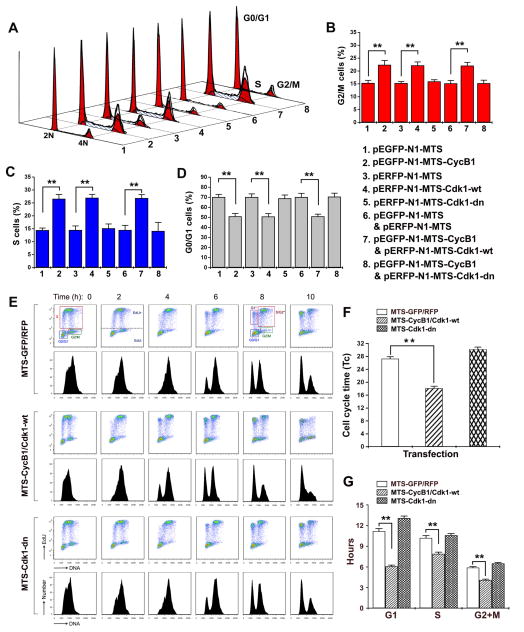
A substantial amount of mitochondrial energy is required for cell-cycle progression. The mechanisms underlying the coordination of the mitochondrial respiration with cell-cycle progression, especially the G2/M transition, remain to be elucidated. Here, we show that a fraction of cyclin B1/Cdk1 proteins localizes to the matrix of mitochondria and phosphorylates a cluster of mitochondrial proteins, including the complex I (CI) subunits in the respiratory chain. Cyclin B1/Cdk1-mediated CI phosphorylation enhances CI activity, whereas deficiency of such phosphorylation in each of the relevant CI subunits results in impairment of CI function. Mitochondria-targeted cyclin B1/Cdk1 increases mitochondrial respiration with enhanced oxygen consumption and ATP generation, which provides cells with efficient bioenergy for G2/M transition and shortens overall cell-cycle time. Thus, cyclin B1/Cdk1-mediated phosphorylation of mitochondrial substrates allows cells to sense and respond to increased energy demand for G2/M transition and, subsequently, to upregulate mitochondrial respiration for successful cell-cycle progression.
Copyright © 2014 Elsevier Inc. All rights reserved.
Figures
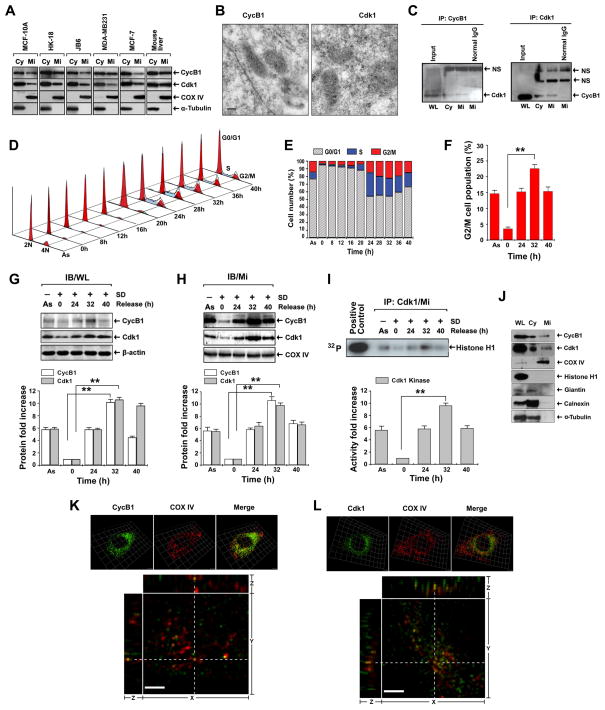
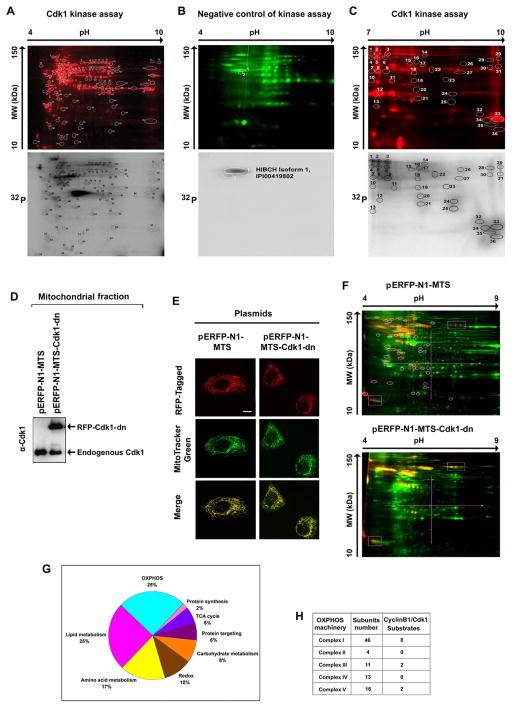
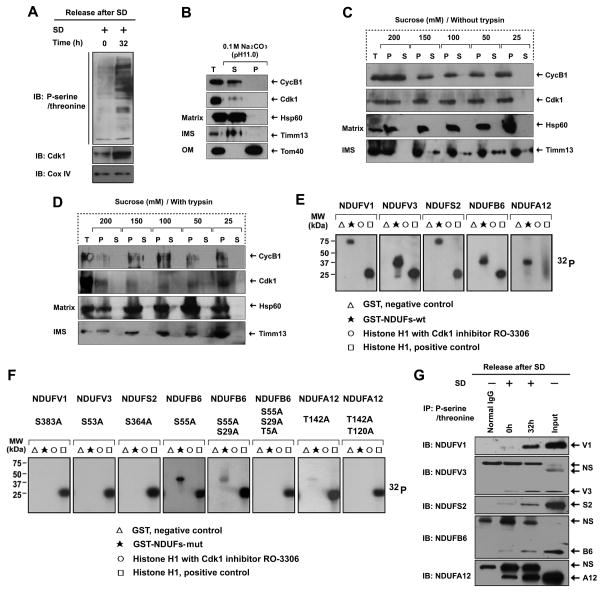
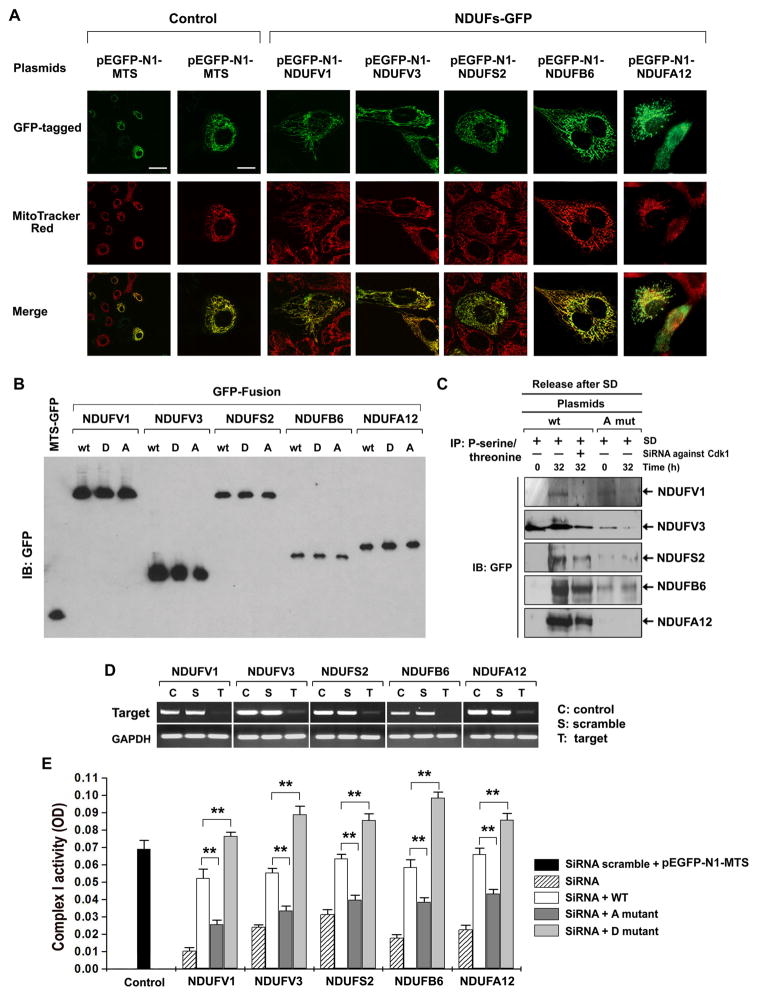
References
-
- Bao Y, Mukai K, Hishiki T, Kubo A, Ohmura M, Sugiura Y, Matsuura T, Nagahata Y, Hayakawa N, Yamamoto T, et al. Energy management by enhanced glycolysis in G1-phase in human colon cancer cells in vitro and in vivo. Mol Cancer Res. 2013;11:973–985. - PubMed
-
- Baumann K. Cell cycle: Order in the pericentriolar material. Nat Rev Mol Cell Biol. 2012;13:749. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
