

Fragile X mental retardation protein regulates translation by binding directly to the ribosome
- PMID: 24746697
- PMCID: PMC4019695
- DOI: 10.1016/j.molcel.2014.03.023
Fragile X mental retardation protein regulates translation by binding directly to the ribosome
Abstract
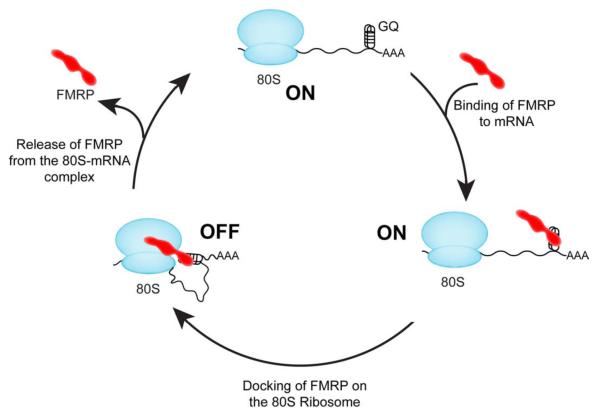
Fragile X syndrome (FXS) is the most common form of inherited mental retardation, and it is caused by loss of function of the fragile X mental retardation protein (FMRP). FMRP is an RNA-binding protein that is involved in the translational regulation of several neuronal mRNAs. However, the precise mechanism of translational inhibition by FMRP is unknown. Here, we show that FMRP inhibits translation by binding directly to the L5 protein on the 80S ribosome. Furthermore, cryoelectron microscopic reconstruction of the 80S ribosome⋅FMRP complex shows that FMRP binds within the intersubunit space of the ribosome such that it would preclude the binding of tRNA and translation elongation factors on the ribosome. These findings suggest that FMRP inhibits translation by blocking the essential components of the translational machinery from binding to the ribosome.
Copyright © 2014 Elsevier Inc. All rights reserved.
Figures
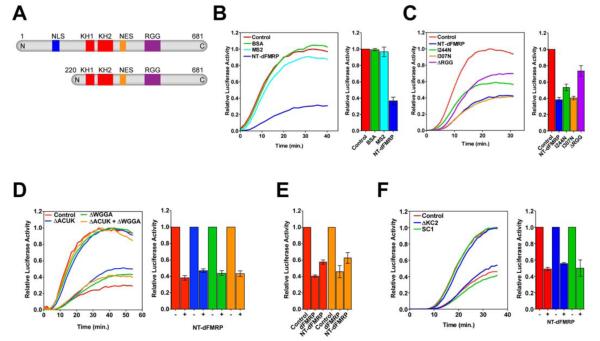
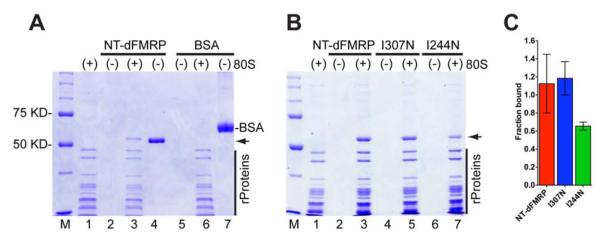
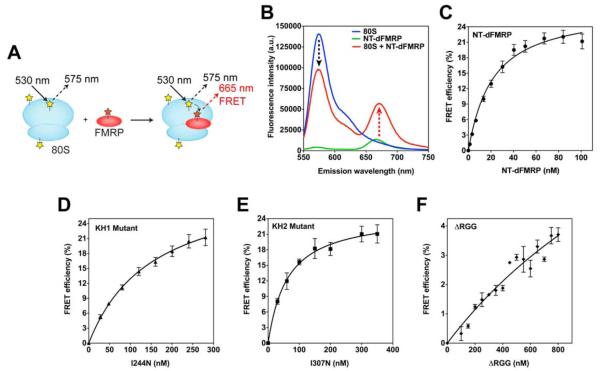
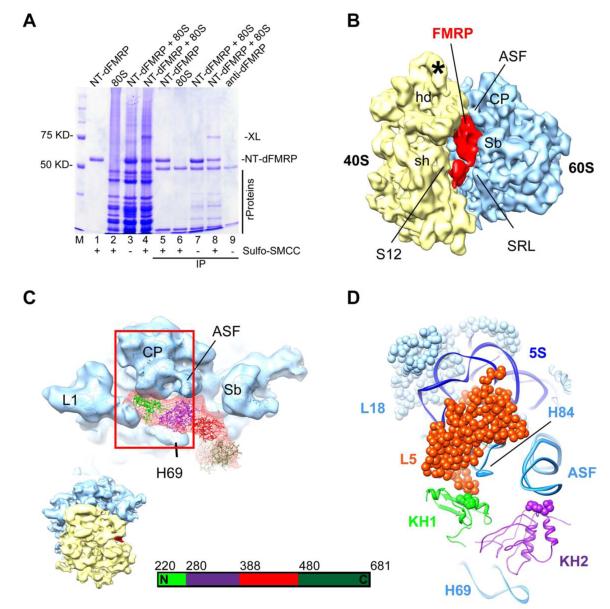
Comment in
-
Fragile X mental retardation protein and the ribosome.Mol Cell. 2014 May 8;54(3):330-2. doi: 10.1016/j.molcel.2014.04.027. Mol Cell. 2014. PMID: 24813710
References
-
- Agulhon C, Blanchet P, Kobetz A, Marchant D, Faucon N, Sarda P, Moraine C, Sittler A, Biancalana V, Malafosse A, et al. Expression of FMR1, FXR1, and FXR2 genes in human prenatal tissues. Journal of neuropathology and experimental neurology. 1999;58:867–880. - PubMed
-
- Ashley CT, Jr., Wilkinson KD, Reines D, Warren ST. FMR1 protein: conserved RNP family domains and selective RNA binding. Science. 1993;262:563–566. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases