

Structural biology-based insights into combinatorial readout and crosstalk among epigenetic marks
- PMID: 24747177
- PMCID: PMC4689310
- DOI: 10.1016/j.bbagrm.2014.04.011
Structural biology-based insights into combinatorial readout and crosstalk among epigenetic marks
Abstract
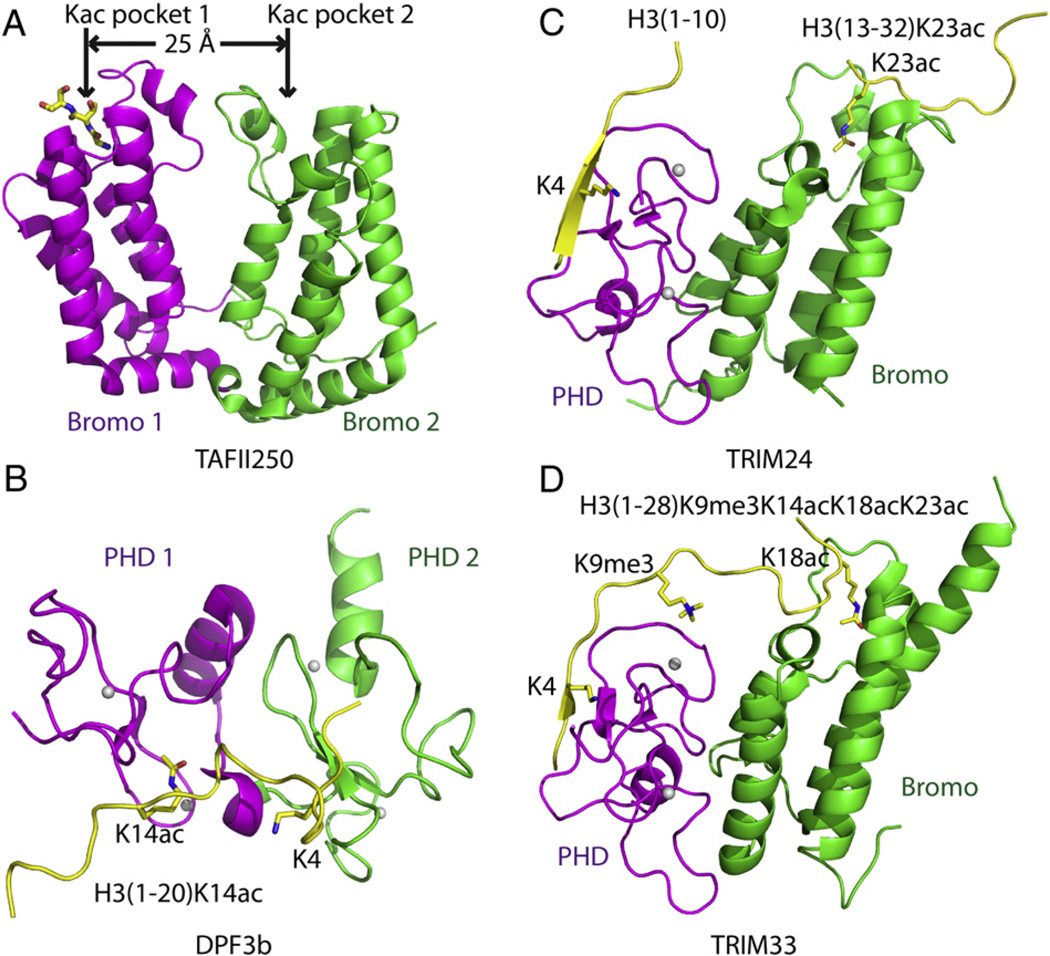
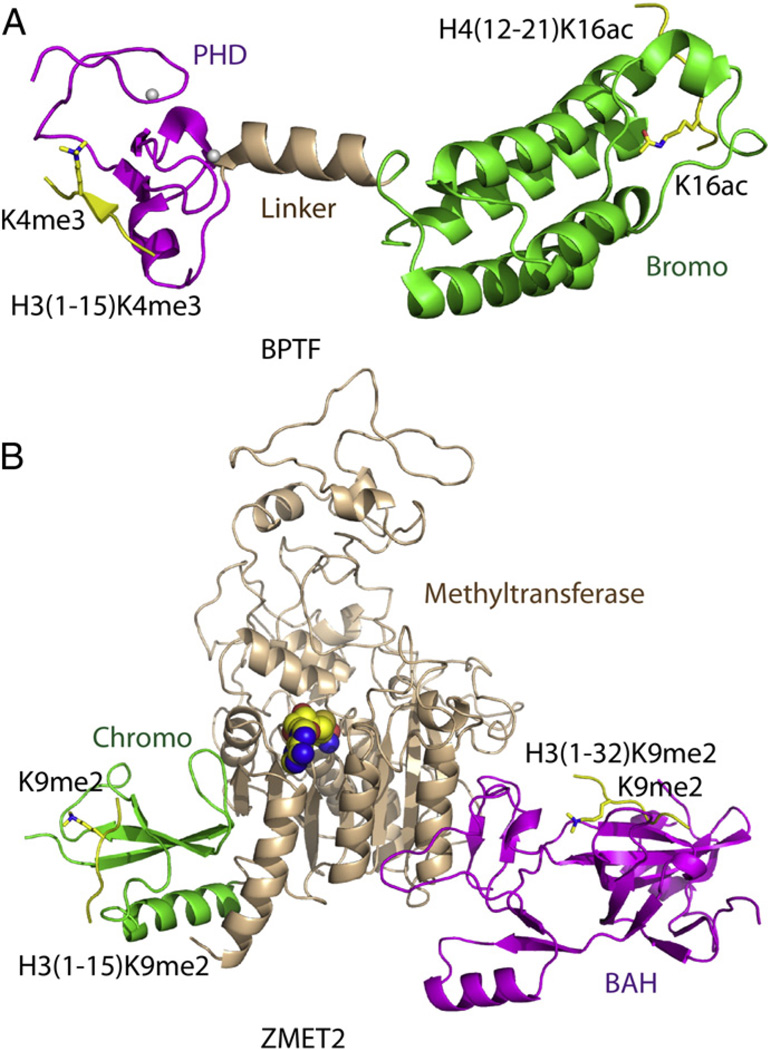
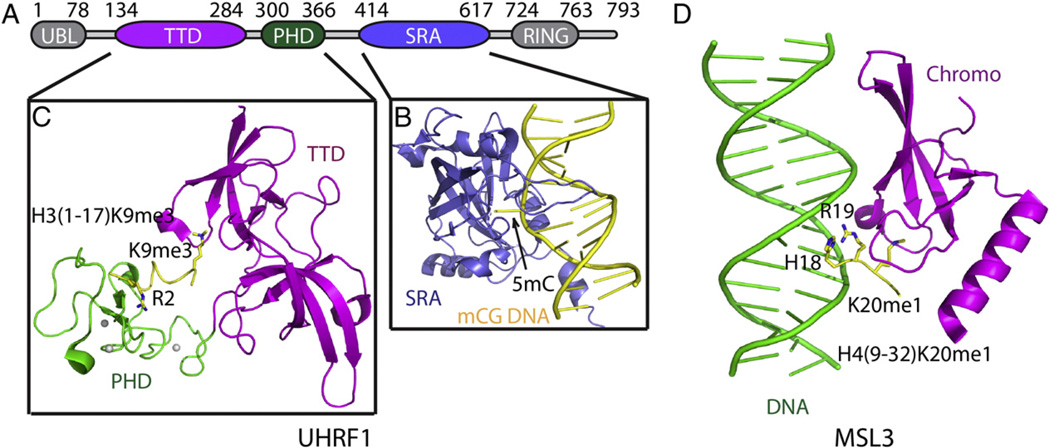
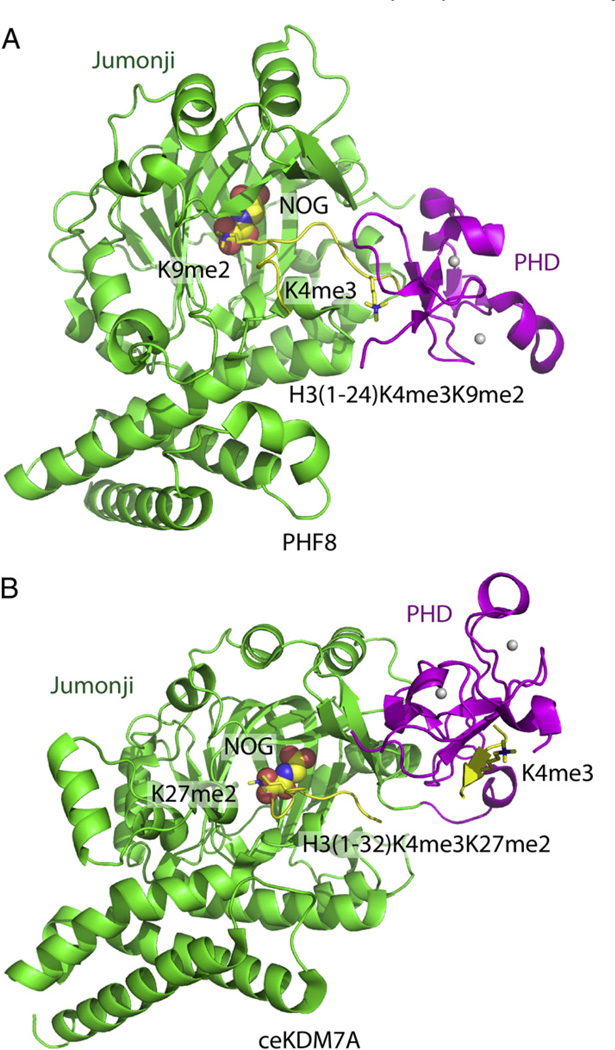
Epigenetic mechanisms control gene regulation by writing, reading and erasing specific epigenetic marks. Within the context of multi-disciplinary approaches applied to investigate epigenetic regulation in diverse systems, structural biology techniques have provided insights at the molecular level of key interactions between upstream regulators and downstream effectors. The early structural efforts focused on studies at the single domain-single mark level have been rapidly extended to research at the multiple domain-multiple mark level, thereby providing additional insights into connections within the complicated epigenetic regulatory network. This review focuses on recent results from structural studies on combinatorial readout and crosstalk among epigenetic marks. It starts with an overview of multiple readout of histone marks associated with both single and dual histone tails, as well as the potential crosstalk between them. Next, this review further expands on the simultaneous readout by epigenetic modules of histone and DNA marks, thereby establishing connections between histone lysine methylation and DNA methylation at the nucleosomal level. Finally, the review discusses the role of pre-existing epigenetic marks in directing the writing/erasing of certain epigenetic marks. This article is part of a Special Issue entitled: Molecular mechanisms of histone modification function.
Keywords: Combinatorial readout; DNA methylation; Epigenetic regulation; Histone modification.
Copyright © 2014 Elsevier B.V. All rights reserved.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
