

Reproductive neuroendocrine dysfunction in polycystic ovary syndrome: insight from animal models
- PMID: 24747343
- PMCID: PMC4175187
- DOI: 10.1016/j.yfrne.2014.04.002
Reproductive neuroendocrine dysfunction in polycystic ovary syndrome: insight from animal models
Abstract
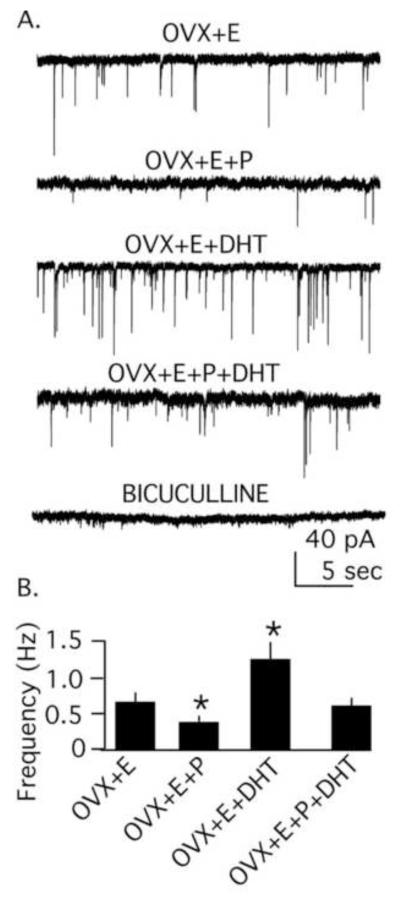
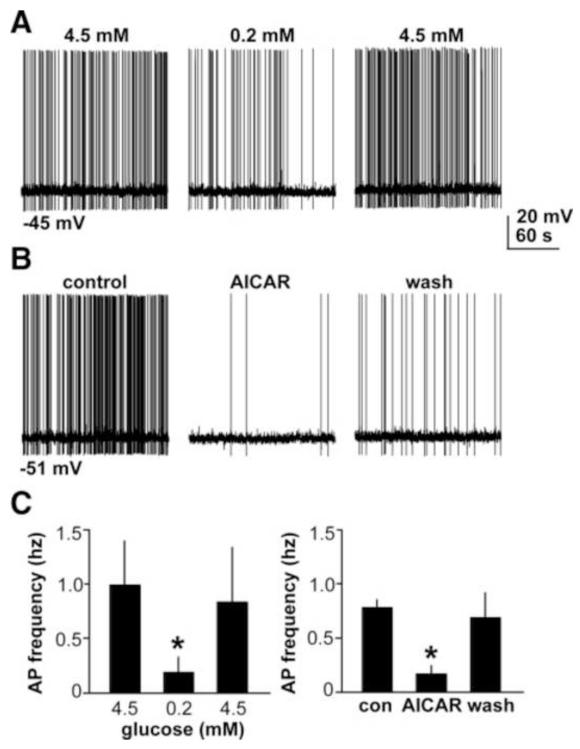
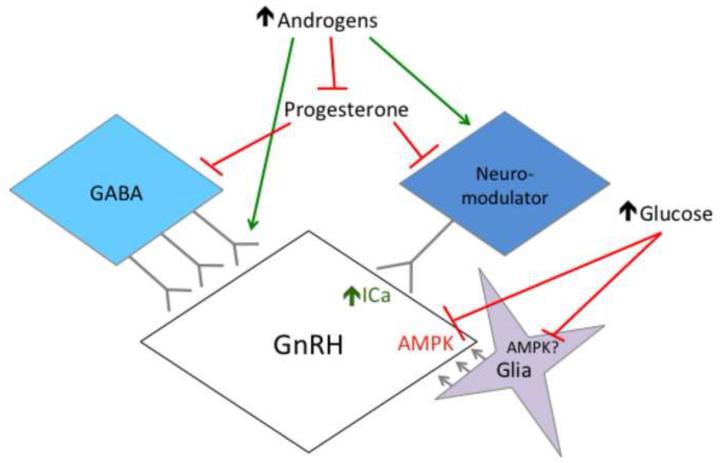
Polycystic ovary syndrome (PCOS) is a common endocrinopathy with elusive origins. A clinically heterogeneous disorder, PCOS is likely to have multiple etiologies comprised of both genetic and environmental factors. Reproductive neuroendocrine dysfunction involving increased frequency and amplitude of gonadotropin-releasing hormone (GnRH) release, as reflected by pulsatile luteinizing hormone (LH) secretion, is an important pathophysiologic component in PCOS. Whether this defect is primary or secondary to other changes in PCOS is unclear, but it contributes significantly to ongoing reproductive dysfunction. This review highlights recent work in animal models, with a particular emphasis on the mouse, demonstrating the ability of pre- and postnatal steroidal and metabolic factors to drive changes in GnRH/LH pulsatility and GnRH neuron function consistent with the observed abnormalities in PCOS. This work has begun to elucidate how a complex interplay of ovarian, metabolic, and neuroendocrine factors culminates in this syndrome.
Keywords: Androgens; Gonadotropin-releasing hormone neurons; Polycystic ovary syndrome; Prenatal androgenization.
Copyright © 2014 Elsevier Inc. All rights reserved.
Figures
References
-
- Abbott DH, Barnett DK, Bruns CM, Dumesic DA. Androgen excess fetal programming of female reproduction: a developmental aetiology for polycystic ovary syndrome? Hum. Reprod. Update. 2005;11:357–374. doi:10.1093/humupd/dmi013. - PubMed
-
- Abbott DH, Bruns CR, Barnett DK, Dunaif A, Goodfriend TL, Dumesic DA, Tarantal AF. Experimentally induced gestational androgen excess disrupts glucoregulation in rhesus monkey dams and their female offspring. Am. J. Physiol. Endocrinol. Metab. 2010;299:E741–751. doi:10.1152/ajpendo.00058.2010. - PMC - PubMed
-
- Abbott DH, Dumesic DA, Eisner JR, Colman RJ, Kemnitz JW. Insights into the development of polycystic ovary syndrome (PCOS) from studies of prenatally androgenized female rhesus monkeys. Trends Endocrinol. Metab. TEM. 1998;9:62–67. - PubMed
-
- Abbott DH, Dumesic DA, Franks S. Developmental origin of polycystic ovary syndrome - a hypothesis. J. Endocrinol. 2002;174:1–5. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
