

The microbiota regulates neutrophil homeostasis and host resistance to Escherichia coli K1 sepsis in neonatal mice
- PMID: 24747744
- PMCID: PMC4016187
- DOI: 10.1038/nm.3542
The microbiota regulates neutrophil homeostasis and host resistance to Escherichia coli K1 sepsis in neonatal mice
Abstract
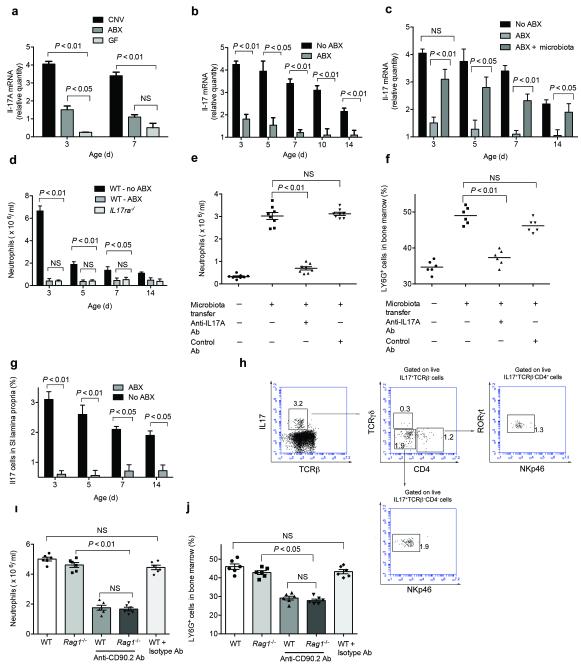
Neonatal colonization by microbes, which begins immediately after birth, is influenced by gestational age and the mother's microbiota and is modified by exposure to antibiotics. In neonates, prolonged duration of antibiotic therapy is associated with increased risk of late-onset sepsis (LOS), a disorder controlled by neutrophils. A role for the microbiota in regulating neutrophil development and susceptibility to sepsis in the neonate remains unclear. We exposed pregnant mouse dams to antibiotics in drinking water to limit transfer of maternal microbes to the neonates. Antibiotic exposure of dams decreased the total number and composition of microbes in the intestine of the neonates. This was associated with decreased numbers of circulating and bone marrow neutrophils and granulocyte/macrophage-restricted progenitor cells in the bone marrow of antibiotic-treated and germ-free neonates. Antibiotic exposure of dams reduced the number of interleukin-17 (IL-17)-producing cells in the intestine and production of granulocyte colony-stimulating factor (G-CSF). Granulocytopenia was associated with impaired host defense and increased susceptibility to Escherichia coli K1 and Klebsiella pneumoniae sepsis in antibiotic-treated neonates, which could be partially reversed by administration of G-CSF. Transfer of a normal microbiota into antibiotic-treated neonates induced IL-17 production by group 3 innate lymphoid cells (ILCs) in the intestine, increasing plasma G-CSF levels and neutrophil numbers in a Toll-like receptor 4 (TLR4)- and myeloid differentiation factor 88 (MyD88)-dependent manner and restored IL-17-dependent resistance to sepsis. Specific depletion of ILCs prevented IL-17- and G-CSF-dependent granulocytosis and resistance to sepsis. These data support a role for the intestinal microbiota in regulation of granulocytosis, neutrophil homeostasis and host resistance to sepsis in neonates.
Figures



Comment in
-
Neonates, antibiotics and the microbiome.Nat Med. 2014 May;20(5):469-70. doi: 10.1038/nm.3558. Nat Med. 2014. PMID: 24804751 No abstract available.
References
-
- Penders J, et al. Factors influencing the composition of the intestinal microbiota in early infancy. Pediatrics. 2006;118:511–521. - PubMed
-
- Sarkar S, Bhagat I, Hieber S, Donn SM. Can neutrophil responses in very low birth weight infants predict the organisms responsible for late-onset bacterial or fungal sepsis? Journal of perinatology: official journal of the California Perinatal Association. 2006;26:501–505. - PubMed
-
- Schwiertz A, et al. Development of the intestinal bacterial composition in hospitalized preterm infants in comparison with breast-fed, full-term infants. Pediatric research. 2003;54:393–399. - PubMed
-
- Pflughoeft KJ, Versalovic J. Human microbiome in health and disease. Annual review of pathology. 2012;7:99–122. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01 AI099479/AI/NIAID NIH HHS/United States
- 3R37HL079142/HL/NHLBI NIH HHS/United States
- R01 HL105834/HL/NHLBI NIH HHS/United States
- 5R01AI038446/AI/NIAID NIH HHS/United States
- 5T32HD060556/HD/NICHD NIH HHS/United States
- R01 HL062052/HL/NHLBI NIH HHS/United States
- 5R01HL105834/HL/NHLBI NIH HHS/United States
- 1R01AI099479/AI/NIAID NIH HHS/United States
- R01 AI105168/AI/NIAID NIH HHS/United States
- R01 AI038446/AI/NIAID NIH HHS/United States
- R37 HL079142/HL/NHLBI NIH HHS/United States
- P60 AA009803/AA/NIAAA NIH HHS/United States
- T32 HD060556/HD/NICHD NIH HHS/United States
- P30 DK034987/DK/NIDDK NIH HHS/United States
- 5R01AI093566/AI/NIAID NIH HHS/United States
- R01 AI093566/AI/NIAID NIH HHS/United States
- 1R01AI105168/AI/NIAID NIH HHS/United States
- 5R01HL062052/HL/NHLBI NIH HHS/United States
- R37 AI038446/AI/NIAID NIH HHS/United States
- 5P60AA009803/AA/NIAAA NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
