

Ubiquilin1 represses migration and epithelial-to-mesenchymal transition of human non-small cell lung cancer cells
- PMID: 24747970
- PMCID: PMC4205225
- DOI: 10.1038/onc.2014.97
Ubiquilin1 represses migration and epithelial-to-mesenchymal transition of human non-small cell lung cancer cells
Abstract
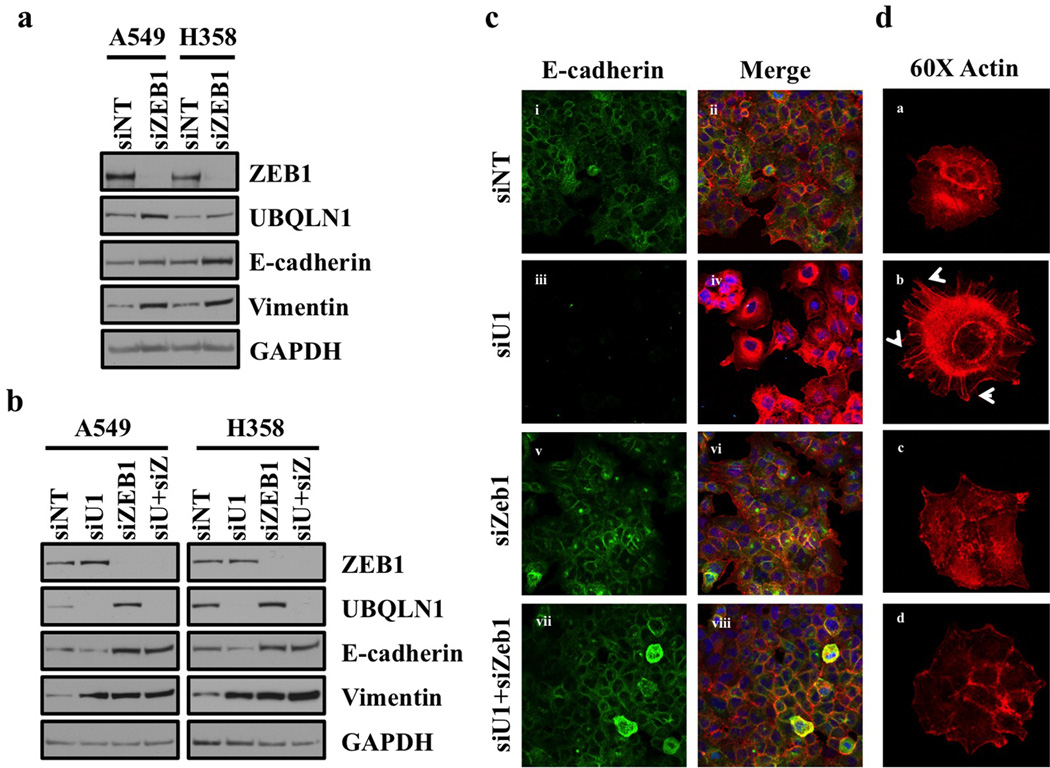
Ubiquilin1 (UBQLN1) is a ubiquitin-like domain and a ubiquitin-associated domain containing protein that has been reported to be involved in shuttling proteins to the proteasome, especially during endoplasmic reticulum-associated protein degradation. Thus, UBQLN1 function has been shown to be critical for combating a number of neurological disorders caused by protein aggregation, such as amyotrophic lateral sclerosis, Alzheimer's disease and Huntington's disease. A role for UBQLN1 in regulating processes involved in tumorigenesis has not been demonstrated. Herein, we show that loss of UBQLN1 causes increased cell migration and invasion, actin cytoskeleton reorganization and induction of epithelial-to-mesenchymal transition (EMT). Loss of UBQLN1 results in a significant decrease in the expression of epithelial markers including E-cadherin and claudin1, whereas expression of mesenchymal markers including Vimentin, Snail and ZEB1 are significantly elevated. Interestingly, we found that ZEB1 is required for induction of mesenchymal-like properties following loss of UBQLN1 and ZEB1 is capable of repressing expression of UBQLN1, suggesting a physiological, reciprocal regulation of EMT by UBQLN1 and ZEB1. Further, we find evidence for a role for UBQLN2 in also regulating EMT and cell migration. These observations have potential clinical relevance because the UBQLN1 gene is lost and underexpressed in a large percentage of human cancer cell lines, and primary human lung cancer samples and recurrent mutations in all five UBQLN family members have been identified in human lung cancers. Taken together, our results suggest for the first time a role for UBQLN family members in cancer biology.
Conflict of interest statement
Figures
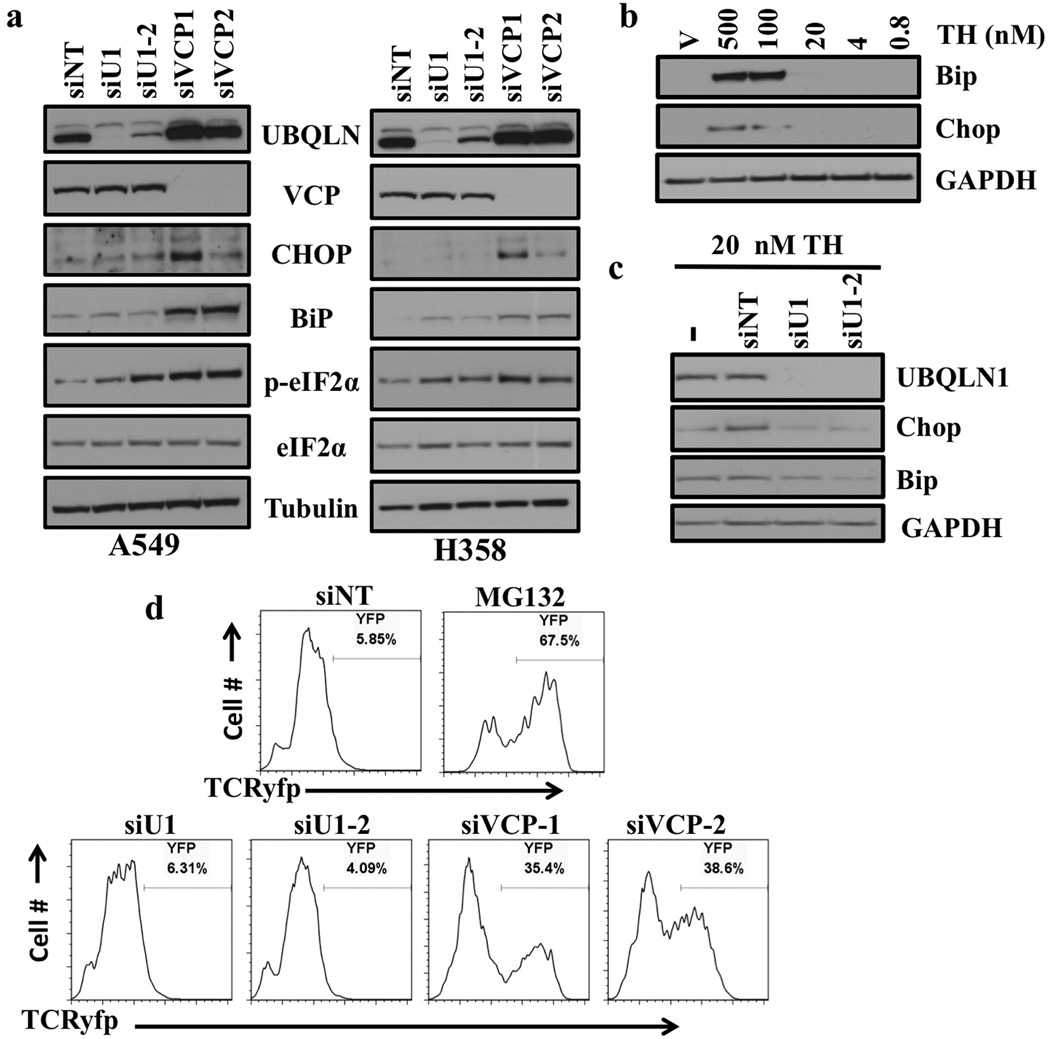
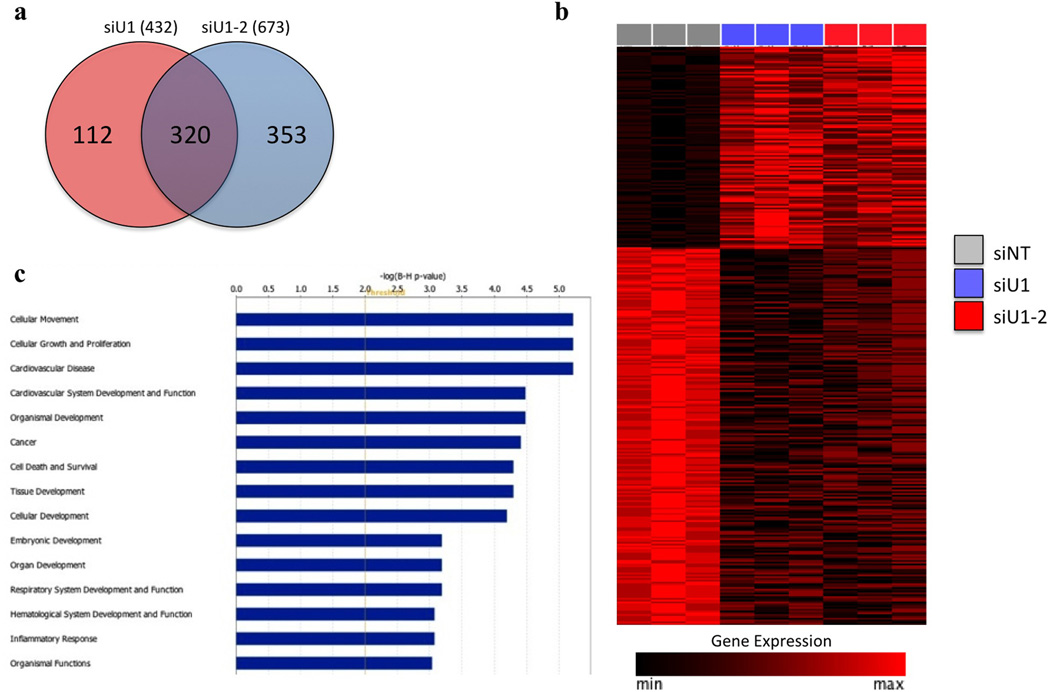
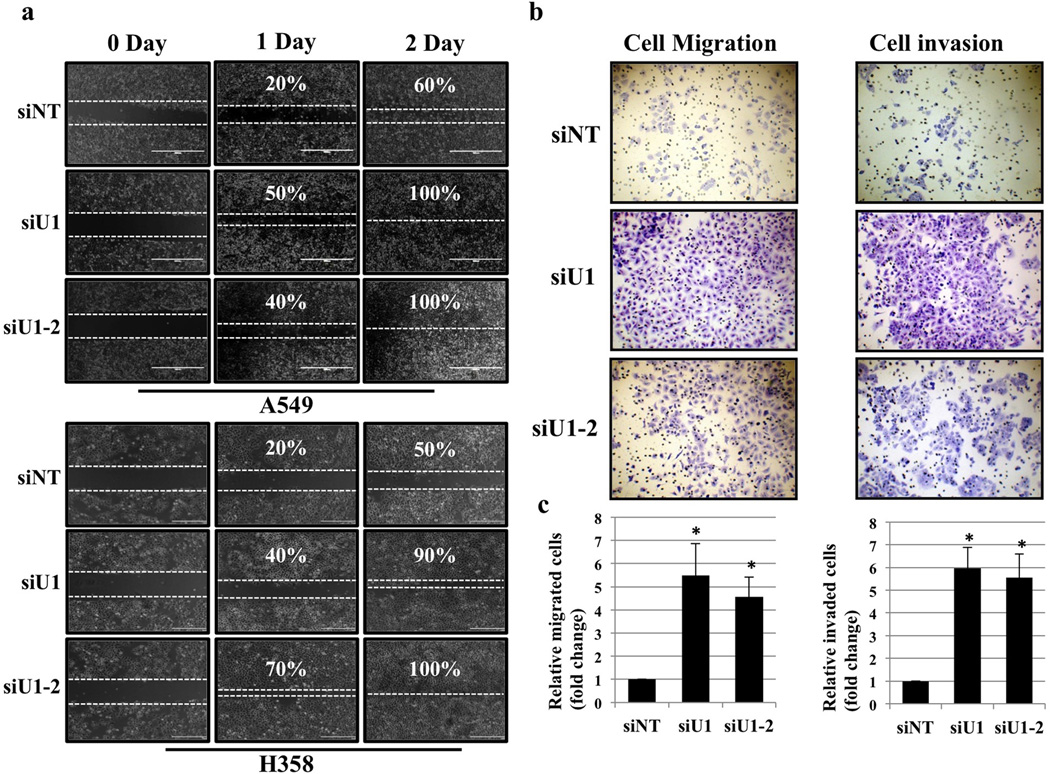
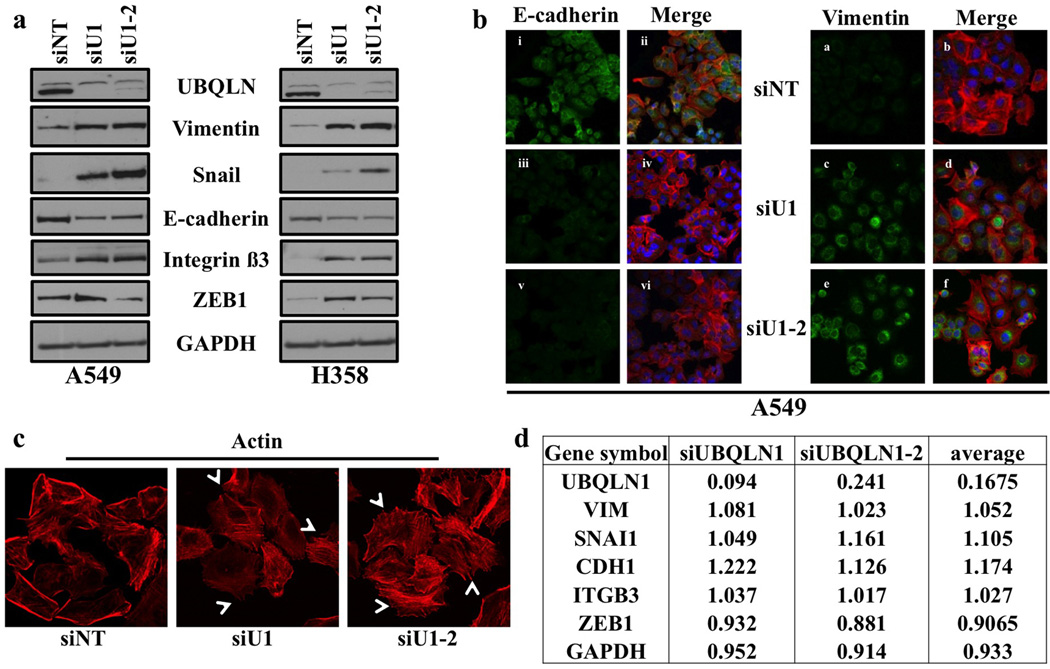


References
-
- Lu A, Hiltunen M, Romano DM, Soininen H, Hyman BT, Bertram L, et al. Effects of ubiquilin 1 on the unfolded protein response. J Mol Neurosci. 2009;38:19–30. - PubMed
-
- Arias-Vásquez A, de Lau L, Pardo L, Liu F, Feng BJ, Bertoli-Avella A, et al. Relationship of the Ubiquilin 1 gene with Alzheimer's and Parkinson's disease and cognitive function. Neurosci Lett. 2007;424:1–5. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
