

Cortical control of adaptation and sensory relay mode in the thalamus
- PMID: 24748112
- PMCID: PMC4020068
- DOI: 10.1073/pnas.1318665111
Cortical control of adaptation and sensory relay mode in the thalamus
Abstract
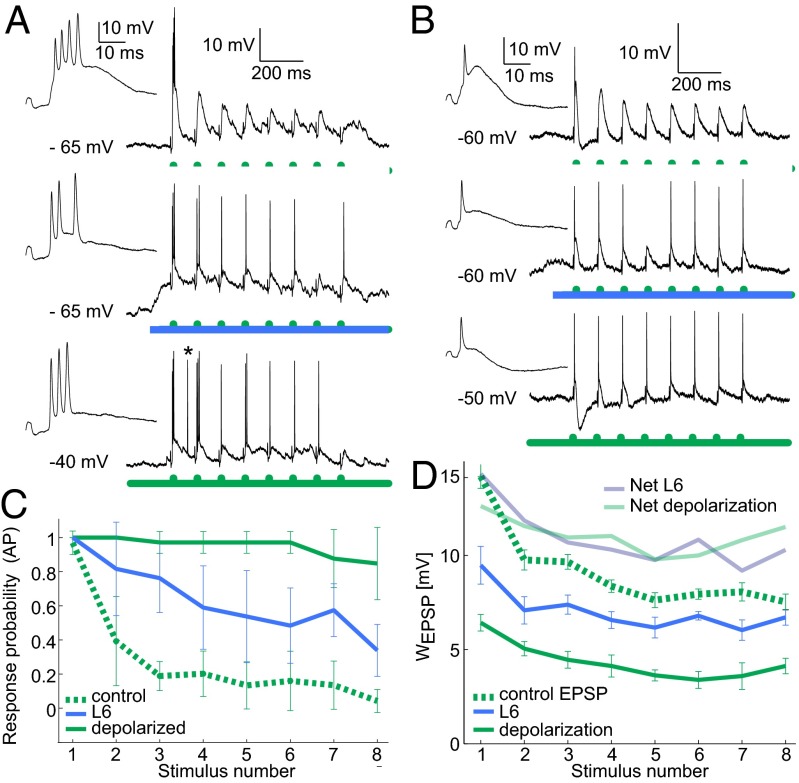
A major synaptic input to the thalamus originates from neurons in cortical layer 6 (L6); however, the function of this cortico-thalamic pathway during sensory processing is not well understood. In the mouse whisker system, we found that optogenetic stimulation of L6 in vivo results in a mixture of hyperpolarization and depolarization in the thalamic target neurons. The hyperpolarization was transient, and for longer L6 activation (>200 ms), thalamic neurons reached a depolarized resting membrane potential which affected key features of thalamic sensory processing. Most importantly, L6 stimulation reduced the adaptation of thalamic responses to repetitive whisker stimulation, thereby allowing thalamic neurons to relay higher frequencies of sensory input. Furthermore, L6 controlled the thalamic response mode by shifting thalamo-cortical transmission from bursting to single spiking. Analysis of intracellular sensory responses suggests that L6 impacts these thalamic properties by controlling the resting membrane potential and the availability of the transient calcium current IT, a hallmark of thalamic excitability. In summary, L6 input to the thalamus can shape both the overall gain and the temporal dynamics of sensory responses that reach the cortex.
Keywords: Ntsr1; firing mode; low threshold calcium spike; sensory systems; top-down modulation.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
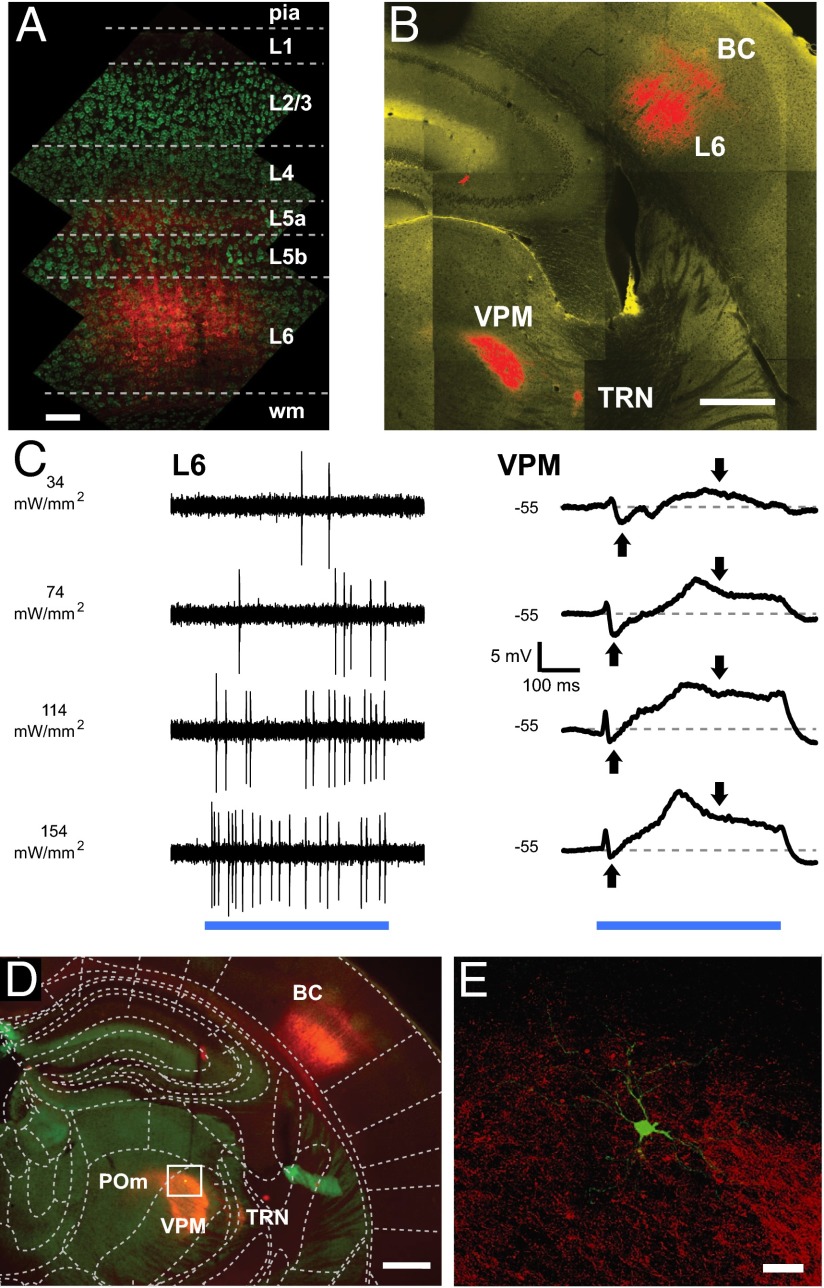
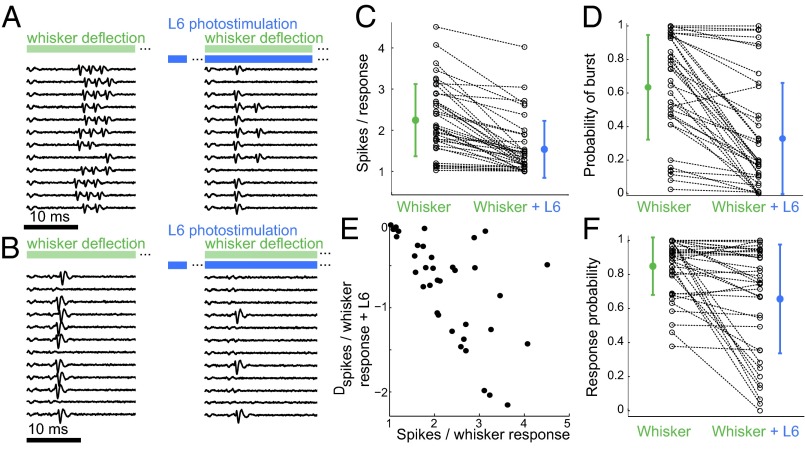
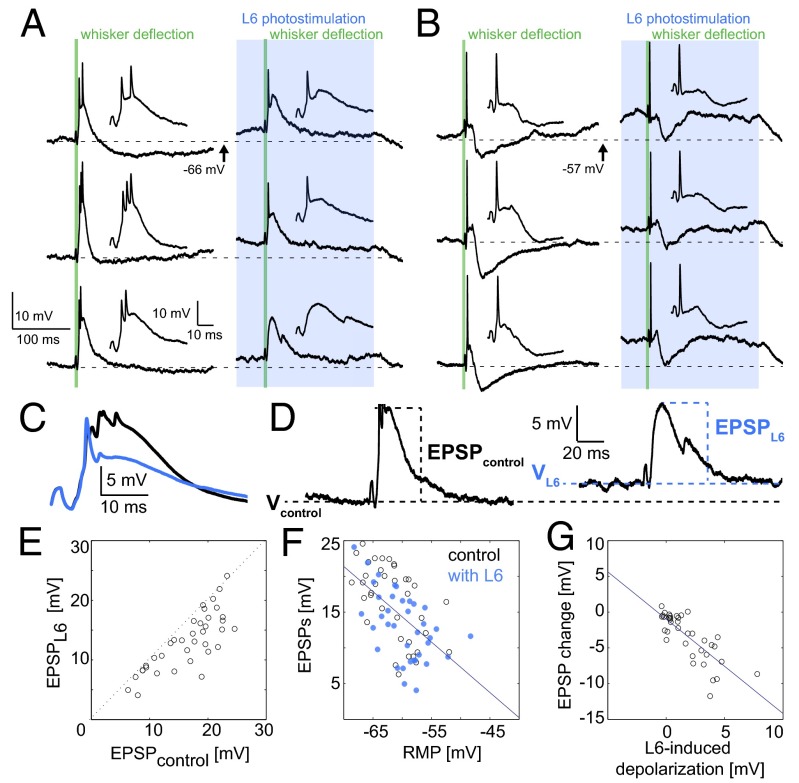
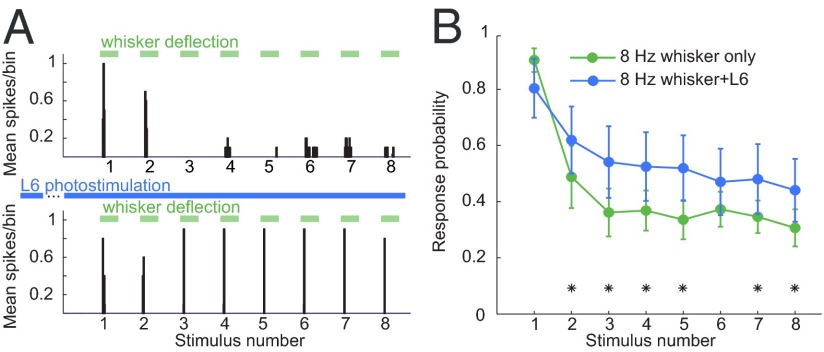
References
-
- Fairhall AL, Lewen GD, Bialek W, de Ruyter Van Steveninck RR. Efficiency and ambiguity in an adaptive neural code. Nature. 2001;412(6849):787–792. - PubMed
-
- Barlow H. Possible Principles Underlying the Transformation of Sensory Messages. Cambridge, MA: MIT Press; 1961.
-
- Diamond ME, Armstrong-James M, Ebner FF. Somatic sensory responses in the rostral sector of the posterior group (POm) and in the ventral posterior medial nucleus (VPM) of the rat thalamus. J Comp Neurol. 1992;318(4):462–476. - PubMed
-
- Simons DJ, Carvell GE. Thalamocortical response transformation in the rat vibrissa/barrel system. J Neurophysiol. 1989;61(2):311–330. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
