

A new lncRNA, APTR, associates with and represses the CDKN1A/p21 promoter by recruiting polycomb proteins
- PMID: 24748121
- PMCID: PMC3991591
- DOI: 10.1371/journal.pone.0095216
A new lncRNA, APTR, associates with and represses the CDKN1A/p21 promoter by recruiting polycomb proteins
Abstract
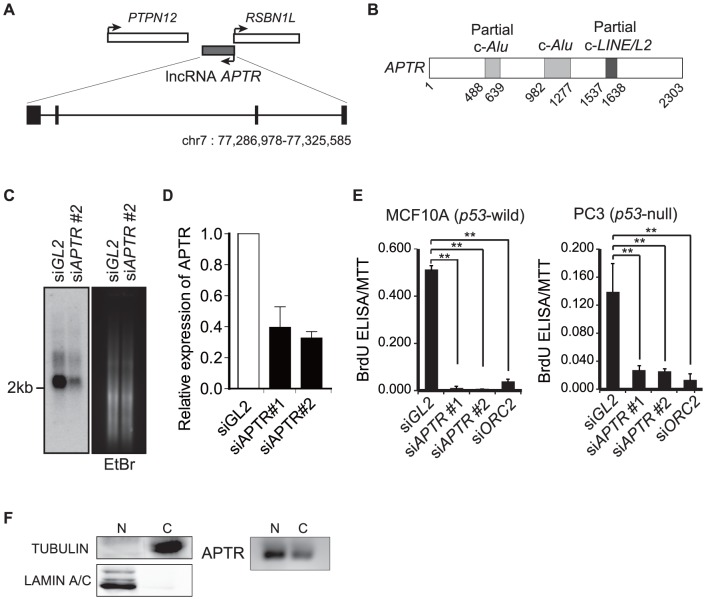
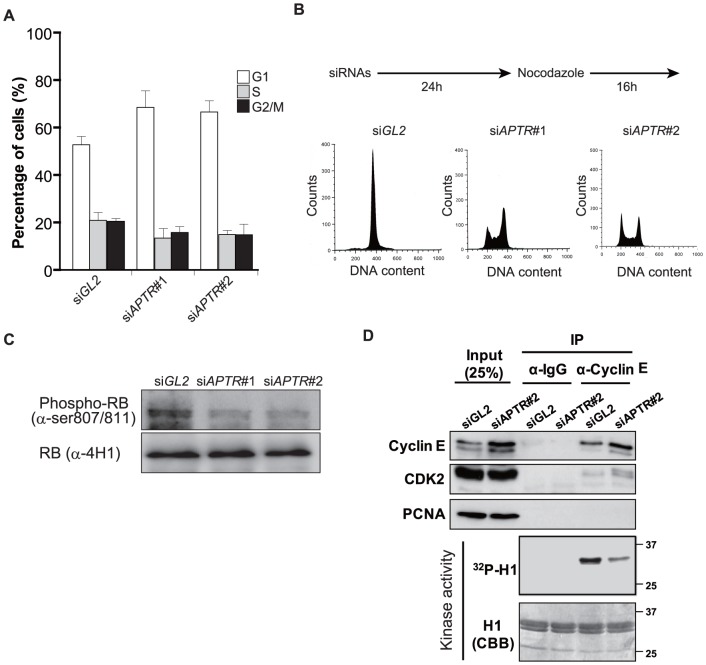
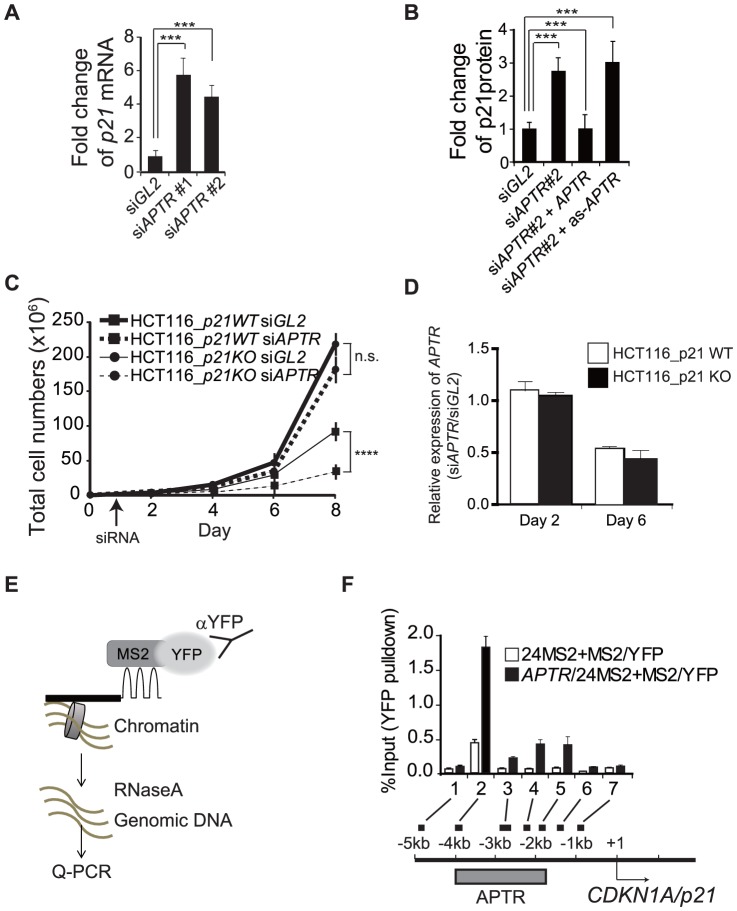
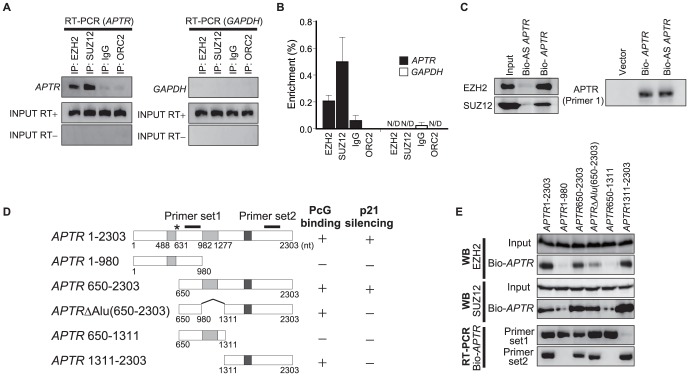
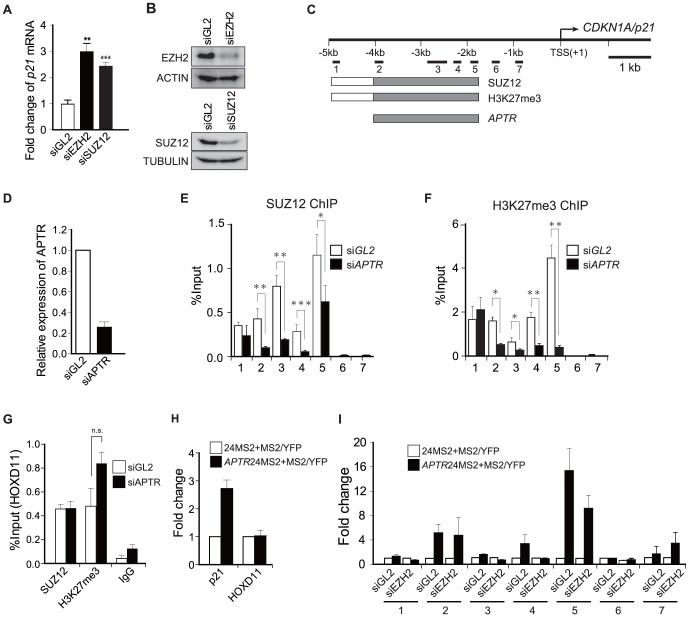
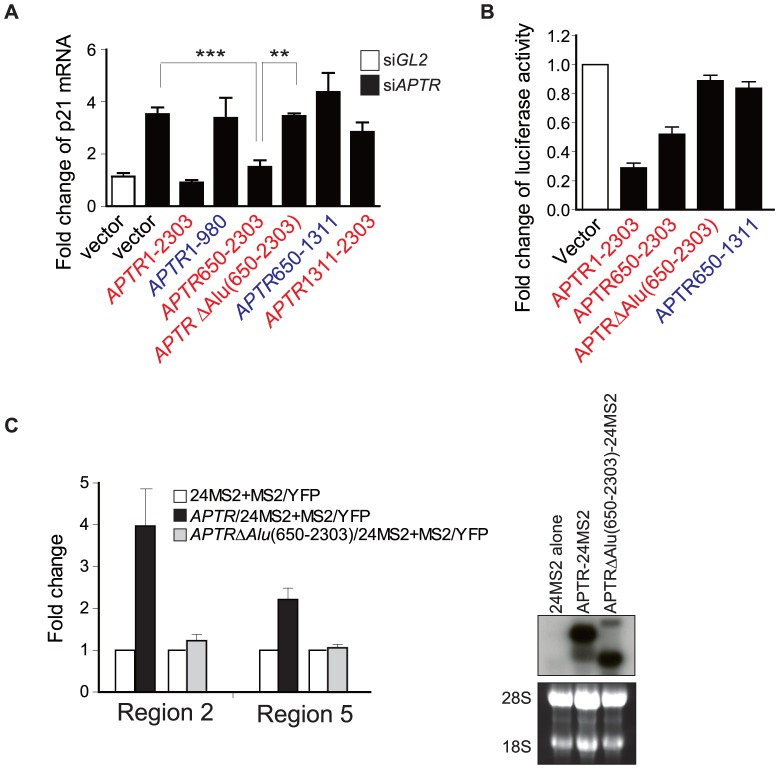
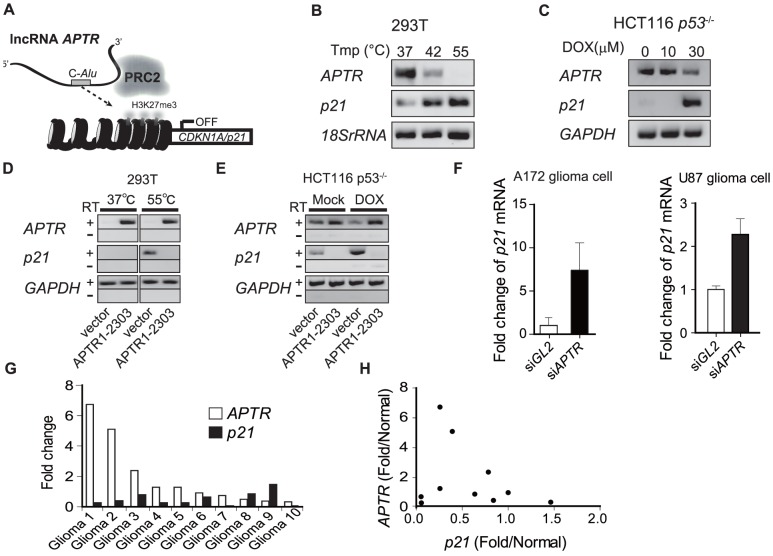
Long noncoding RNAs (lncRNAs) have emerged as a major regulator of cell physiology, but many of which have no known function. CDKN1A/p21 is an important inhibitor of the cell-cycle, regulator of the DNA damage response and effector of the tumor suppressor p53, playing a crucial role in tumor development and prevention. In order to identify a regulator for tumor progression, we performed an siRNA screen of human lncRNAs required for cell proliferation, and identified a novel lncRNA, APTR, that acts in trans to repress the CDKN1A/p21 promoter independent of p53 to promote cell proliferation. APTR associates with the promoter of CDKN1A/p21 and this association requires a complementary-Alu sequence encoded in APTR. A different module of APTR associates with and recruits the Polycomb repressive complex 2 (PRC2) to epigenetically repress the p21 promoter. A decrease in APTR is necessary for the induction of p21 after heat stress and DNA damage by doxorubicin, and the levels of APTR and p21 are anti-correlated in human glioblastomas. Our data identify a new regulator of the cell-cycle inhibitor CDKN1A/p21 that acts as a proliferative factor in cancer cell lines and in glioblastomas and demonstrate that Alu elements present in lncRNAs can contribute to targeting regulatory lncRNAs to promoters.
Conflict of interest statement
Figures
Similar articles
-
Long noncoding RNA APTR contributes to osteosarcoma progression through repression of miR-132-3p and upregulation of yes-associated protein 1.J Cell Physiol. 2019 Jun;234(6):8998-9007. doi: 10.1002/jcp.27572. Epub 2018 Oct 14. J Cell Physiol. 2019. PMID: 30317613
-
The intronic long noncoding RNA ANRASSF1 recruits PRC2 to the RASSF1A promoter, reducing the expression of RASSF1A and increasing cell proliferation.PLoS Genet. 2013;9(8):e1003705. doi: 10.1371/journal.pgen.1003705. Epub 2013 Aug 22. PLoS Genet. 2013. PMID: 23990798 Free PMC article.
-
A novel long noncoding RNA Linc-ASEN represses cellular senescence through multileveled reduction of p21 expression.Cell Death Differ. 2020 Jun;27(6):1844-1861. doi: 10.1038/s41418-019-0467-6. Epub 2019 Dec 9. Cell Death Differ. 2020. PMID: 31819156 Free PMC article.
-
Long non-coding RNA ANRIL in gene regulation and its duality in atherosclerosis.J Huazhong Univ Sci Technolog Med Sci. 2017 Dec;37(6):816-822. doi: 10.1007/s11596-017-1812-y. Epub 2017 Dec 21. J Huazhong Univ Sci Technolog Med Sci. 2017. PMID: 29270737 Review.
-
The control of polycomb repressive complexes by long noncoding RNAs.Wiley Interdiscip Rev RNA. 2021 Nov;12(6):e1657. doi: 10.1002/wrna.1657. Epub 2021 Apr 16. Wiley Interdiscip Rev RNA. 2021. PMID: 33861025 Free PMC article. Review.
Cited by
-
The Importance of Networking: Plant Polycomb Repressive Complex 2 and Its Interactors.Epigenomes. 2022 Mar 3;6(1):8. doi: 10.3390/epigenomes6010008. Epigenomes. 2022. PMID: 35323212 Free PMC article. Review.
-
Vernalization-Triggered Intragenic Chromatin Loop Formation by Long Noncoding RNAs.Dev Cell. 2017 Feb 6;40(3):302-312.e4. doi: 10.1016/j.devcel.2016.12.021. Epub 2017 Jan 26. Dev Cell. 2017. PMID: 28132848 Free PMC article.
-
LncRNAs Act as a Link between Chronic Liver Disease and Hepatocellular Carcinoma.Int J Mol Sci. 2020 Apr 20;21(8):2883. doi: 10.3390/ijms21082883. Int J Mol Sci. 2020. PMID: 32326098 Free PMC article. Review.
-
Long Noncoding RNAs and Human Liver Disease.Annu Rev Pathol. 2022 Jan 24;17:1-21. doi: 10.1146/annurev-pathol-042320-115255. Epub 2021 Aug 20. Annu Rev Pathol. 2022. PMID: 34416820 Free PMC article. Review.
-
LncRNA APTR amplification serves as a potential glioma biomarker and promotes glioma progression via miR-6734-5p/ TCF7/LEF1 axis.Noncoding RNA Res. 2025 Feb 22;12:42-55. doi: 10.1016/j.ncrna.2025.02.007. eCollection 2025 Jun. Noncoding RNA Res. 2025. PMID: 40103614 Free PMC article.
References
-
- Bracken AP, Helin K (2009) Polycomb group proteins: navigators of lineage pathways led astray in cancer. Nat Rev Cancer 9: 773–784. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
