

Genome-wide RNA-binding analysis of the trypanosome U1 snRNP proteins U1C and U1-70K reveals cis/trans-spliceosomal network
- PMID: 24748659
- PMCID: PMC4041458
- DOI: 10.1093/nar/gku286
Genome-wide RNA-binding analysis of the trypanosome U1 snRNP proteins U1C and U1-70K reveals cis/trans-spliceosomal network
Abstract
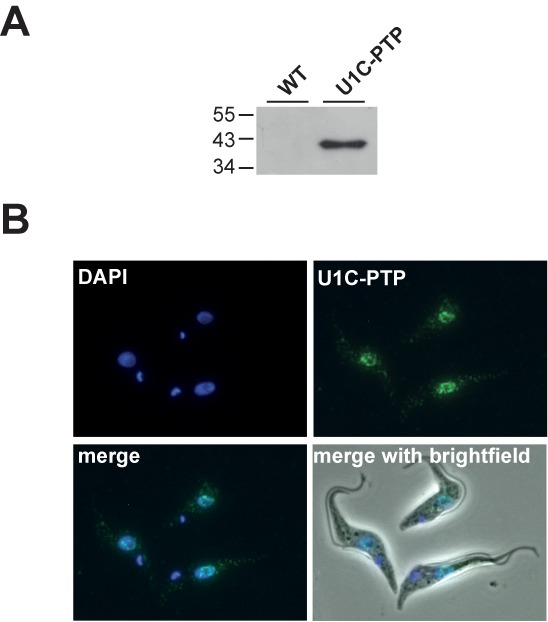
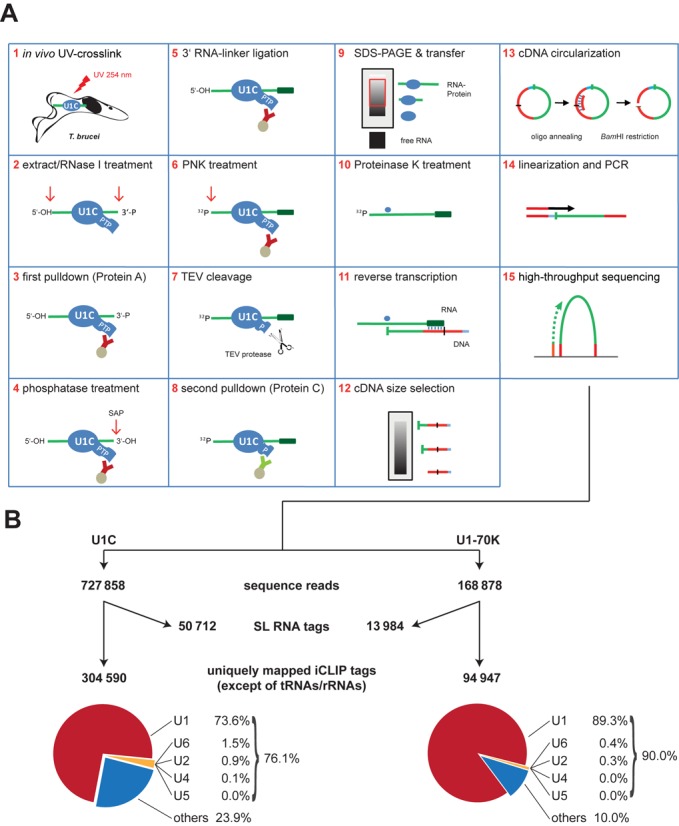
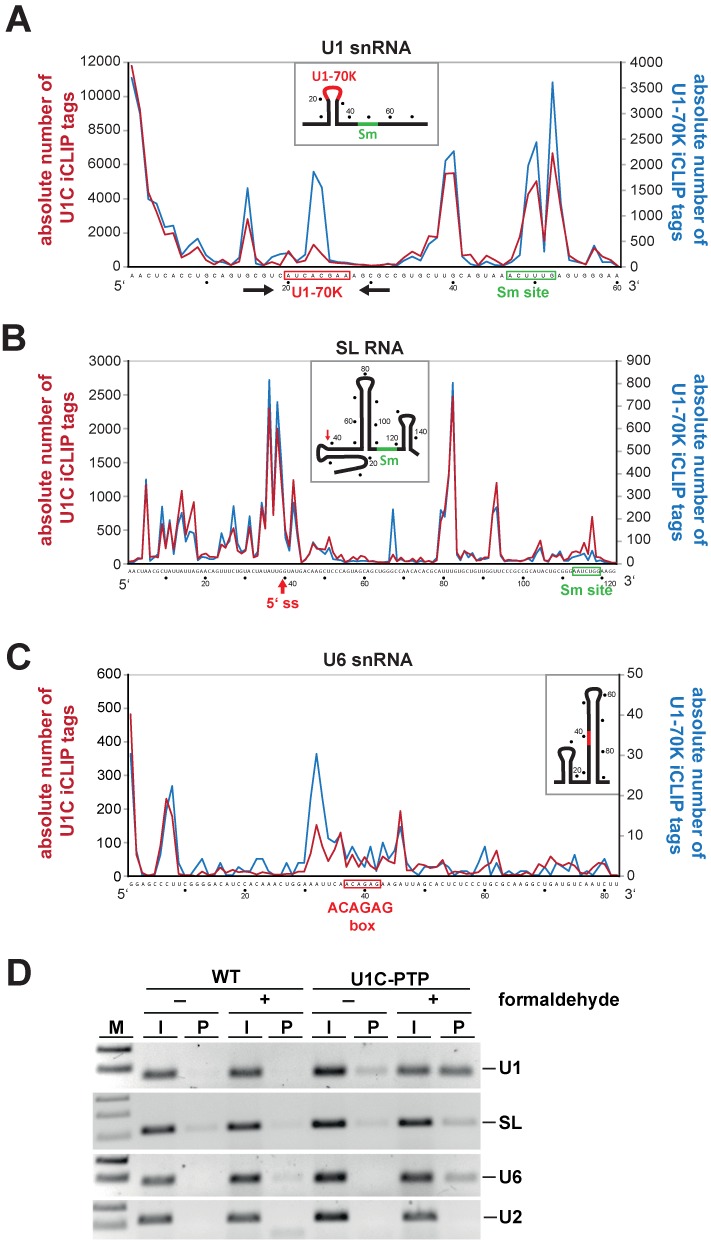
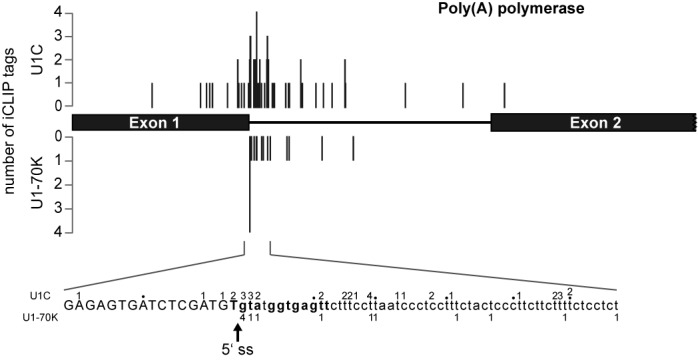
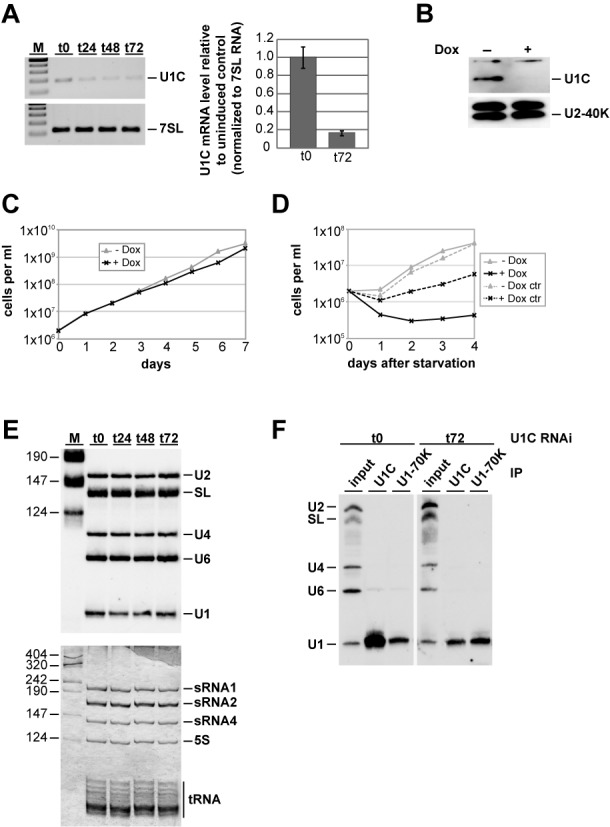
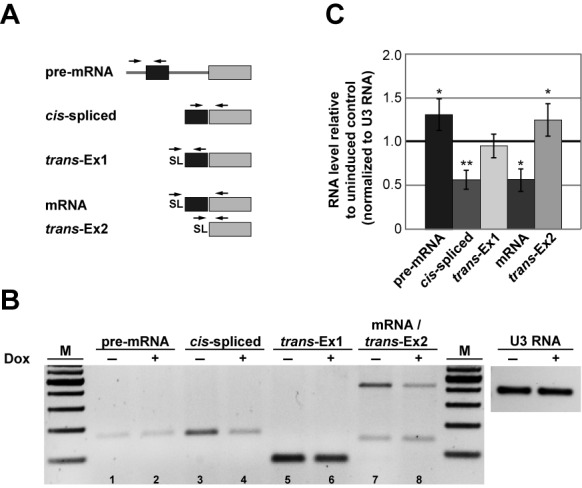
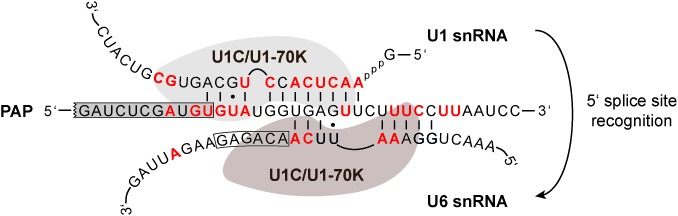
Trans-splicing in trypanosomes adds a 39-nucleotide mini-exon from the spliced leader (SL) RNA to the 5' end of each protein-coding sequence. On the other hand, cis-splicing of the few intron-containing genes requires the U1 small nuclear ribonucleoprotein (snRNP) particle. To search for potential new functions of the U1 snRNP in Trypanosoma brucei, we applied genome-wide individual-nucleotide resolution crosslinking-immunoprecipitation (iCLIP), focusing on the U1 snRNP-specific proteins U1C and U1-70K. Surprisingly, U1C and U1-70K interact not only with the U1, but also with U6 and SL RNAs. In addition, mapping of crosslinks to the cis-spliced PAP [poly(A) polymerase] pre-mRNA indicate an active role of these proteins in 5' splice site recognition. In sum, our results demonstrate that the iCLIP approach provides insight into stable and transient RNA-protein contacts within the spliceosomal network. We propose that the U1 snRNP may represent an evolutionary link between the cis- and trans-splicing machineries, playing a dual role in 5' splice site recognition on the trans-spliceosomal SL RNP as well as on pre-mRNA cis-introns.
© The Author(s) 2014. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures
Similar articles
-
Biochemical and functional characterization of the cis-spliceosomal U1 small nuclear RNP from Trypanosoma brucei.Mol Biochem Parasitol. 2002 May;121(2):233-43. doi: 10.1016/s0166-6851(02)00044-0. Mol Biochem Parasitol. 2002. PMID: 12034457
-
U1 small nuclear RNP from Trypanosoma brucei: a minimal U1 snRNA with unusual protein components.Nucleic Acids Res. 2005 Apr 29;33(8):2493-503. doi: 10.1093/nar/gki548. Print 2005. Nucleic Acids Res. 2005. PMID: 15863726 Free PMC article.
-
Spliceosomal proteomics in Trypanosoma brucei reveal new RNA splicing factors.Eukaryot Cell. 2009 Jul;8(7):990-1000. doi: 10.1128/EC.00075-09. Epub 2009 May 8. Eukaryot Cell. 2009. PMID: 19429779 Free PMC article.
-
Principles and correction of 5'-splice site selection.RNA Biol. 2022 Jan;19(1):943-960. doi: 10.1080/15476286.2022.2100971. RNA Biol. 2022. PMID: 35866748 Free PMC article. Review.
-
A novel role of U1 snRNP: Splice site selection from a distance.Biochim Biophys Acta Gene Regul Mech. 2019 Jun;1862(6):634-642. doi: 10.1016/j.bbagrm.2019.04.004. Epub 2019 Apr 28. Biochim Biophys Acta Gene Regul Mech. 2019. PMID: 31042550 Free PMC article. Review.
Cited by
-
Regulation of gene expression in trypanosomatids: living with polycistronic transcription.Open Biol. 2019 Jun 28;9(6):190072. doi: 10.1098/rsob.190072. Epub 2019 Jun 5. Open Biol. 2019. PMID: 31164043 Free PMC article.
-
Computational analysis of sense-antisense chimeric transcripts reveals their potential regulatory features and the landscape of expression in human cells.NAR Genom Bioinform. 2021 Aug 25;3(3):lqab074. doi: 10.1093/nargab/lqab074. eCollection 2021 Sep. NAR Genom Bioinform. 2021. PMID: 34458728 Free PMC article.
-
The polyadenylation complex of Trypanosoma brucei: Characterization of the functional poly(A) polymerase.RNA Biol. 2016;13(2):221-31. doi: 10.1080/15476286.2015.1130208. RNA Biol. 2016. PMID: 26727667 Free PMC article.
-
SNRPC promotes hepatocellular carcinoma cell motility by inducing epithelial-mesenchymal transition.FEBS Open Bio. 2021 Jun;11(6):1757-1770. doi: 10.1002/2211-5463.13175. Epub 2021 May 12. FEBS Open Bio. 2021. PMID: 33934562 Free PMC article.
-
Evolution and stress response potential of the plant splicing factor U1C.Sci Rep. 2024 Jul 26;14(1):17212. doi: 10.1038/s41598-024-68190-4. Sci Rep. 2024. PMID: 39060315 Free PMC article.
References
-
- Preußer C., Jaé N., Günzl A., Bindereif A. Pre-mRNA splicing in Trypanosoma brucei: factors, mechanisms, and regulation. In: Bindereif A, editor. mRNA Metabolism in Trypanosomes. Berlin-Heidelberg: Springer-Verlag GmbH; 2012. pp. 49–77.
-
- Berriman M., Ghedin E., Hertz-Fowler C., Blandin G., Renauld H., Bartholomeu D.C., Lennard N.J., Caler E., Hamlin N.E., Haas B., et al. The genome of the African trypanosome Trypanosoma brucei. Science. 2005;309:416–422. - PubMed
-
- Djikeng A., Ferreira L., D’Angelo M., Dolezal P., Lamb T., Murta S., Triggs V., Ulbert S., Villarino A., Renzi S., et al. Characterization of a candidate Trypanosoma brucei U1 small nuclear RNA gene. Mol. Biochem. Parasitol. 2001;113:109–115. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
