

Structural and biophysical characterization of Staphylococcus aureus SaMazF shows conservation of functional dynamics
- PMID: 24748664
- PMCID: PMC4041440
- DOI: 10.1093/nar/gku266
Structural and biophysical characterization of Staphylococcus aureus SaMazF shows conservation of functional dynamics
Abstract
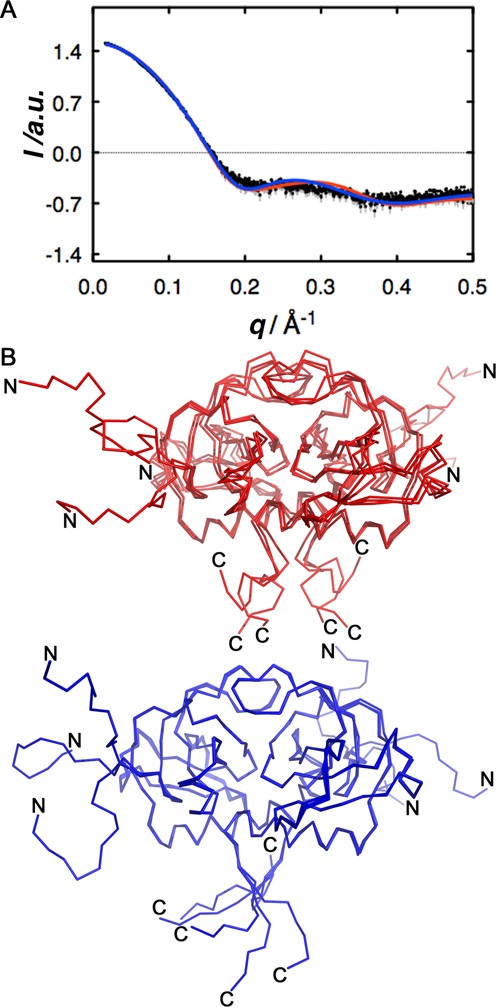
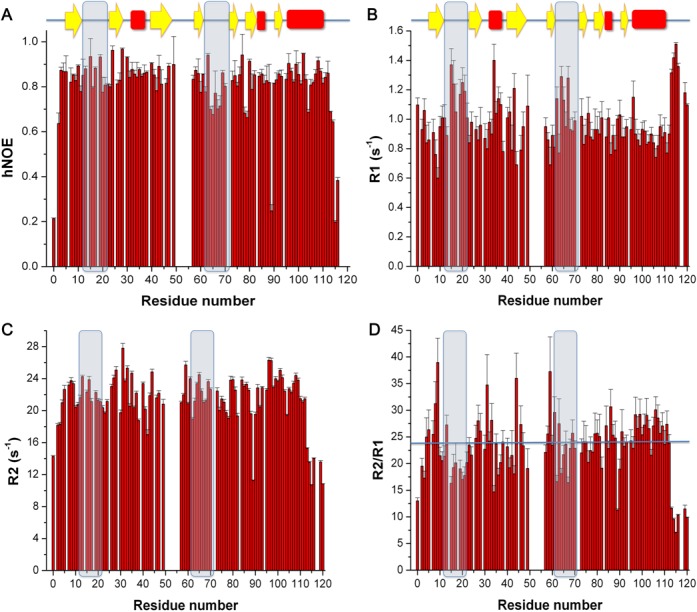
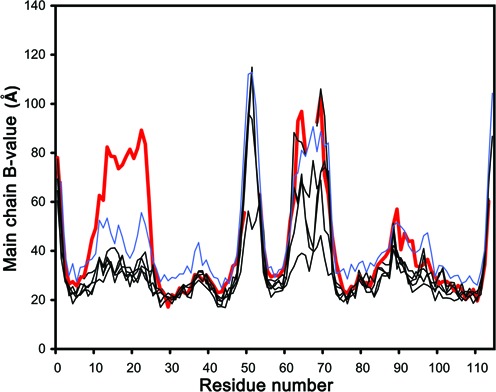
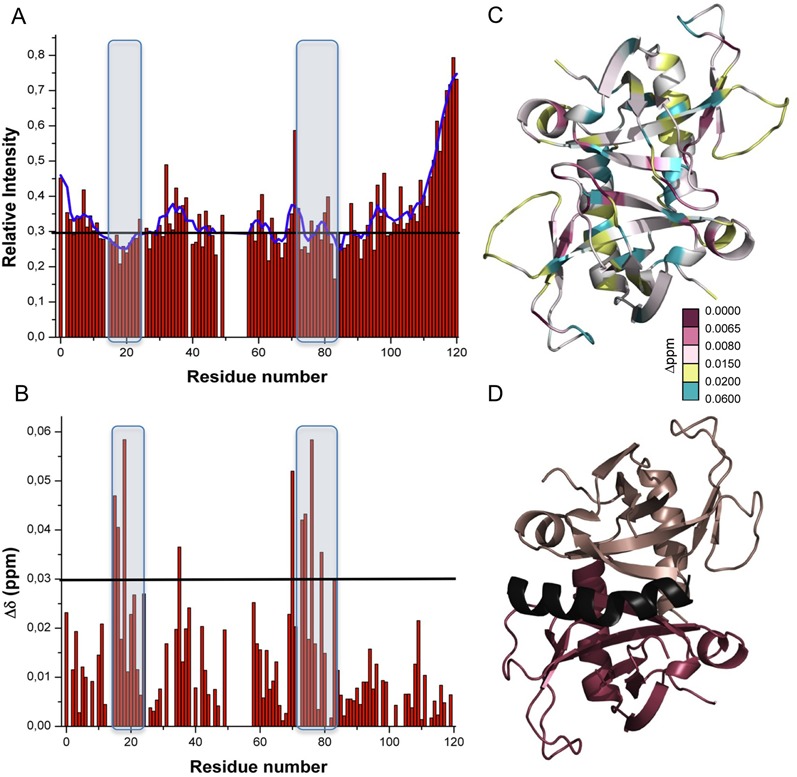
The Staphylococcus aureus genome contains three toxin-antitoxin modules, including one mazEF module, SamazEF. Using an on-column separation protocol we are able to obtain large amounts of wild-type SaMazF toxin. The protein is well-folded and highly resistant against thermal unfolding but aggregates at elevated temperatures. Crystallographic and nuclear magnetic resonance (NMR) solution studies show a well-defined dimer. Differences in structure and dynamics between the X-ray and NMR structural ensembles are found in three loop regions, two of which undergo motions that are of functional relevance. The same segments also show functionally relevant dynamics in the distantly related CcdB family despite divergence of function. NMR chemical shift mapping and analysis of residue conservation in the MazF family suggests a conserved mode for the inhibition of MazF by MazE.
© The Author(s) 2014. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures


Similar articles
-
Crystallization of the Staphylococcus aureus MazF mRNA interferase.Acta Crystallogr Sect F Struct Biol Cryst Commun. 2011 Mar 1;67(Pt 3):386-9. doi: 10.1107/S1744309111000571. Epub 2011 Feb 25. Acta Crystallogr Sect F Struct Biol Cryst Commun. 2011. PMID: 21393849 Free PMC article.
-
Structural analyses of the MazEF4 toxin-antitoxin pair in Mycobacterium tuberculosis provide evidence for a unique extracellular death factor.J Biol Chem. 2017 Nov 17;292(46):18832-18847. doi: 10.1074/jbc.M117.807974. Epub 2017 Oct 2. J Biol Chem. 2017. PMID: 28972145 Free PMC article.
-
mazEF: a chromosomal toxin-antitoxin module that triggers programmed cell death in bacteria.J Cell Sci. 2005 Oct 1;118(Pt 19):4327-32. doi: 10.1242/jcs.02619. J Cell Sci. 2005. PMID: 16179604
-
Structure and function of bacterial kid-kis and related toxin-antitoxin systems.Protein Pept Lett. 2007;14(2):113-24. doi: 10.2174/092986607779816096. Protein Pept Lett. 2007. PMID: 17305597 Review.
-
Ribonucleases in bacterial toxin-antitoxin systems.Biochim Biophys Acta. 2013 Jun-Jul;1829(6-7):523-31. doi: 10.1016/j.bbagrm.2013.02.007. Epub 2013 Feb 21. Biochim Biophys Acta. 2013. PMID: 23454553 Review.
Cited by
-
Structural and mutational analysis of MazE6-operator DNA complex provide insights into autoregulation of toxin-antitoxin systems.Commun Biol. 2022 Sep 15;5(1):963. doi: 10.1038/s42003-022-03933-5. Commun Biol. 2022. PMID: 36109664 Free PMC article.
-
MazEF Toxin-Antitoxin System-Mediated DNA Damage Stress Response in Deinococcus radiodurans.Front Genet. 2021 Feb 19;12:632423. doi: 10.3389/fgene.2021.632423. eCollection 2021. Front Genet. 2021. PMID: 33679894 Free PMC article.
-
Role of PemI in the Staphylococcus aureus PemIK toxin-antitoxin complex: PemI controls PemK by acting as a PemK loop mimic.Nucleic Acids Res. 2022 Feb 28;50(4):2319-2333. doi: 10.1093/nar/gkab1288. Nucleic Acids Res. 2022. PMID: 35141752 Free PMC article.
-
Combining cysteine scanning with chemical labeling to map protein-protein interactions and infer bound structure in an intrinsically disordered region.Front Mol Biosci. 2022 Oct 7;9:997653. doi: 10.3389/fmolb.2022.997653. eCollection 2022. Front Mol Biosci. 2022. PMID: 36275627 Free PMC article.
-
Structural and functional analysis of the Klebsiella pneumoniae MazEF toxin-antitoxin system.IUCrJ. 2021 Mar 5;8(Pt 3):362-371. doi: 10.1107/S2052252521000452. eCollection 2021 May 1. IUCrJ. 2021. PMID: 33953923 Free PMC article.
References
-
- Buts L., Lah J., Dao-Thi M., Wyns L., Loris R. Toxin-antitoxin modules as bacterial metabolic stress managers. Trends Biochem. Sci. 2005;30:672–679. - PubMed
-
- Gerdes K., Christensen S.K., Lobner-Olesen A. Prokaryotic toxin-antitoxin stress response loci. Nat. Rev. Microbiol. 2005;3:371–382. - PubMed
-
- Yamaguchi Y., Park J.H., Inouye M. Toxin-antitoxin systems in bacteria and archaea. Annu. Rev. Genet. 2011;45:61–79. - PubMed
-
- Hayes F., Van Melderen L. Toxins-antitoxins: diversity, evolution and function. Crit. Rev. Biochem. Mol. Biol. 2011;46:386–408. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
