

Progenitor cell dynamics in the Newt Telencephalon during homeostasis and neuronal regeneration
- PMID: 24749074
- PMCID: PMC3986684
- DOI: 10.1016/j.stemcr.2014.01.018
Progenitor cell dynamics in the Newt Telencephalon during homeostasis and neuronal regeneration
Abstract
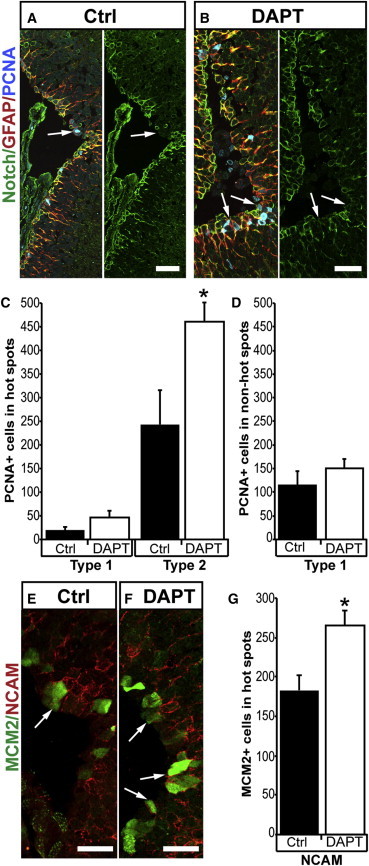
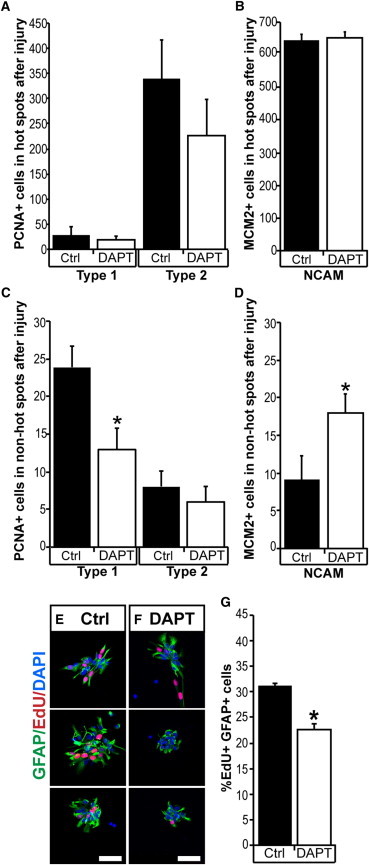
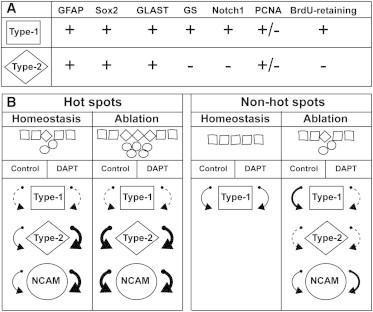
The adult newt brain has a marked neurogenic potential and is highly regenerative. Ventricular, radial glia-like ependymoglia cells give rise to neurons both during normal homeostasis and after injury, but subpopulations among ependymoglia cells have not been defined. We show here that a substantial portion of GFAP(+) ependymoglia cells in the proliferative hot spots of the telencephalon has transit-amplifying characteristics. In contrast, proliferating ependymoglia cells, which are scattered along the ventricular wall, have stem cell features in terms of label retention and insensitivity to AraC treatment. Ablation of neurons remodels the proliferation dynamics and leads to de novo formation of regions displaying features of neurogenic niches, such as the appearance of cells with transit-amplifying features and proliferating neuroblasts. The results have implication both for our understanding of the evolutionary diversification of radial glia cells as well as the processes regulating neurogenesis and regeneration in the adult vertebrate brain.
Figures
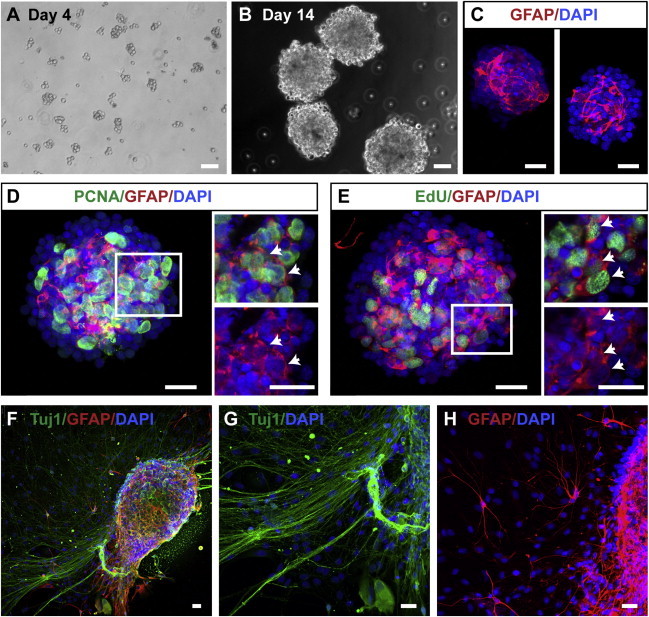
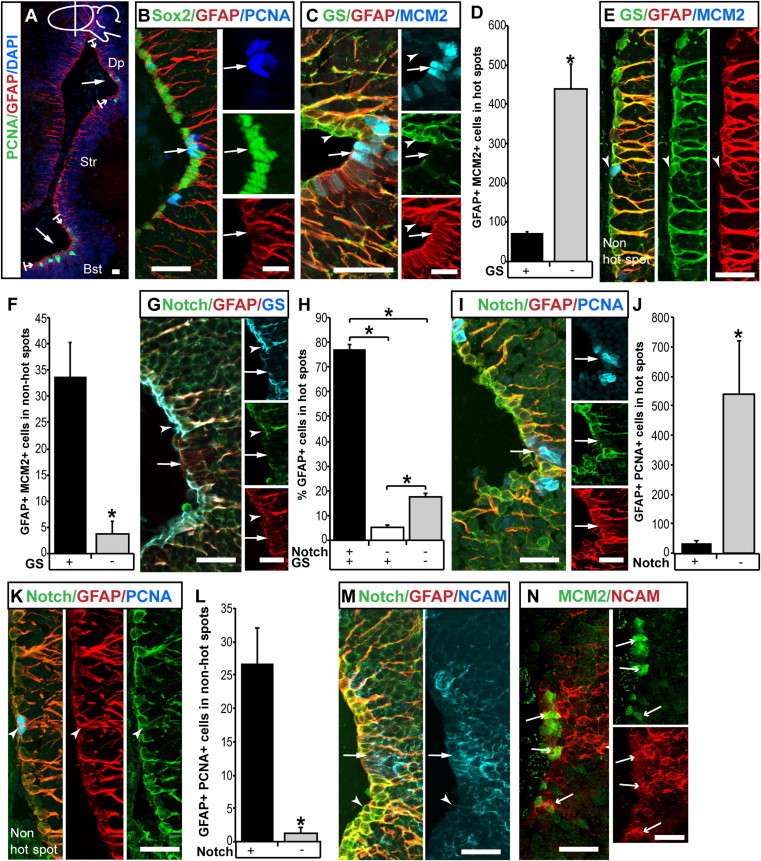
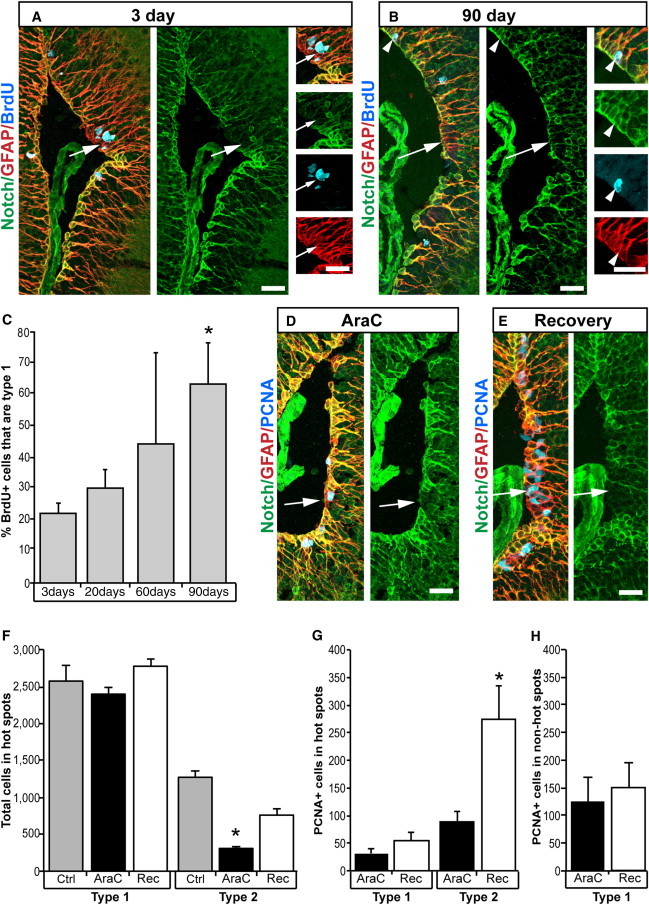
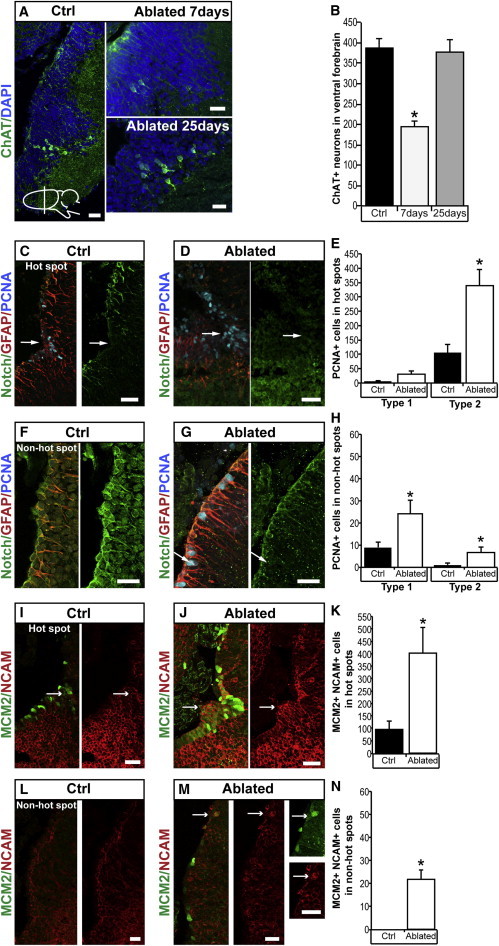
References
-
- Abdullayev I., Kirkham M., Björklund Å.K., Simon A., Sandberg R. A reference transcriptome and inferred proteome for the salamander Notophthalmus viridescens. Exp. Cell Res. 2013;319:1187–1197. - PubMed
-
- Adolf B., Chapouton P., Lam C.S., Topp S., Tannhäuser B., Strähle U., Götz M., Bally-Cuif L. Conserved and acquired features of adult neurogenesis in the zebrafish telencephalon. Dev. Biol. 2006;295:278–293. - PubMed
-
- Arias-Carrión O., Freundlieb N., Oertel W.H., Höglinger G.U. Adult neurogenesis and Parkinson’s disease. CNS Neurol. Disord. Drug Targets. 2007;6:326–335. - PubMed
-
- Arias-Carrión O., Yamada E., Freundlieb N., Djufri M., Maurer L., Hermanns G., Ipach B., Chiu W.-H., Steiner C., Oertel W.H., Höglinger G.U. Neurogenesis in substantia nigra of parkinsonian brains? J. Neural Transm. Suppl. 2009:279–285. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
