

Vinculin phosphorylation differentially regulates mechanotransduction at cell-cell and cell-matrix adhesions
- PMID: 24751539
- PMCID: PMC4003237
- DOI: 10.1083/jcb.201309092
Vinculin phosphorylation differentially regulates mechanotransduction at cell-cell and cell-matrix adhesions
Abstract
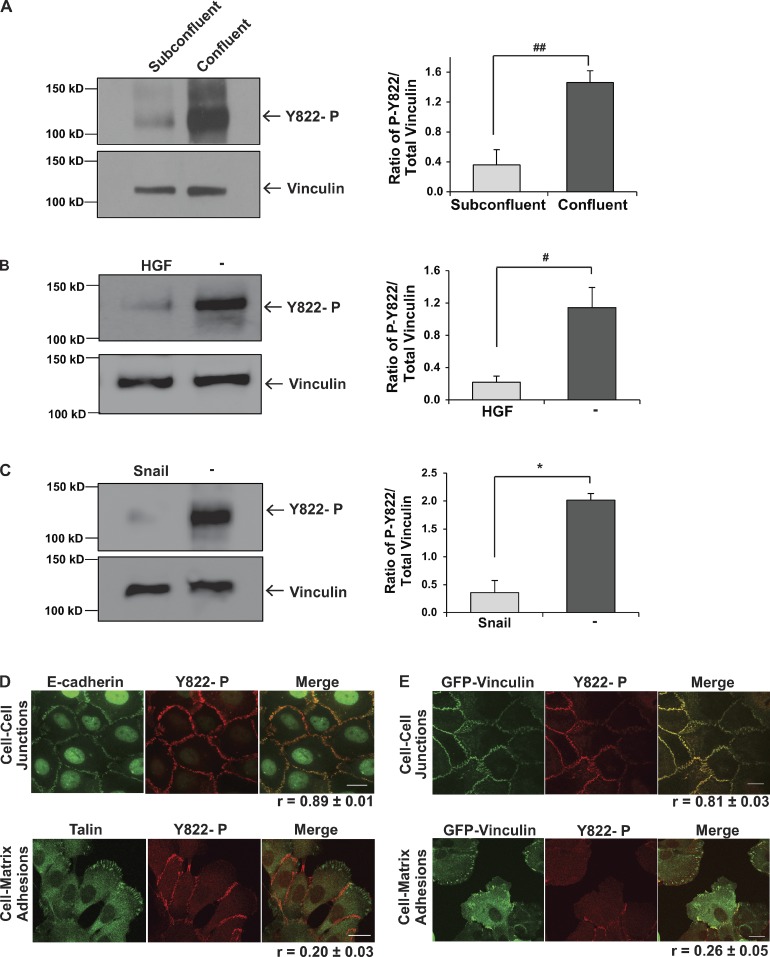
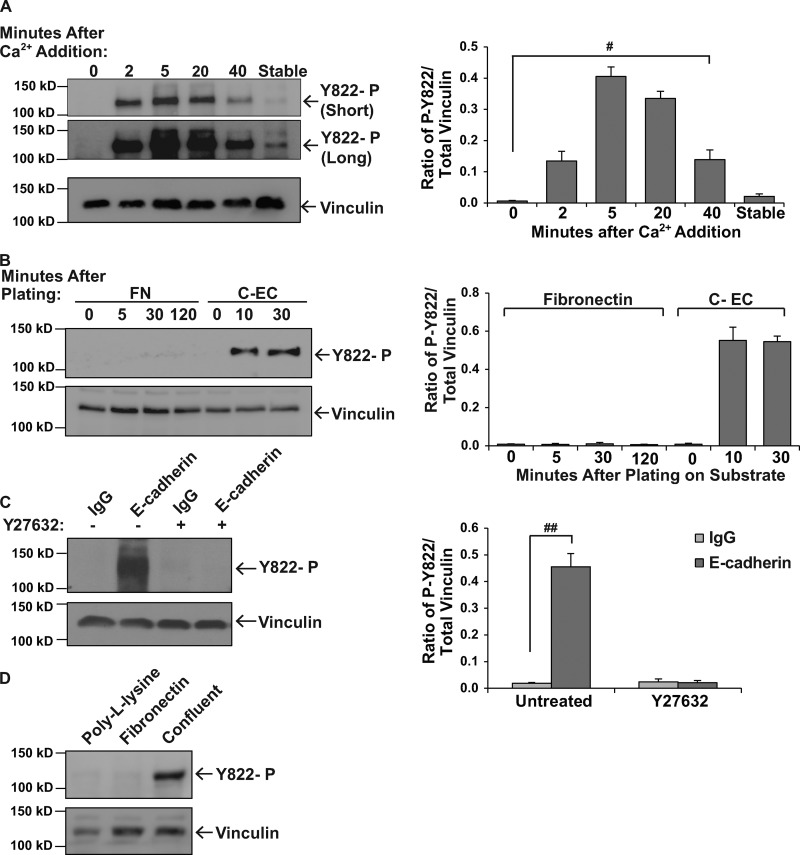
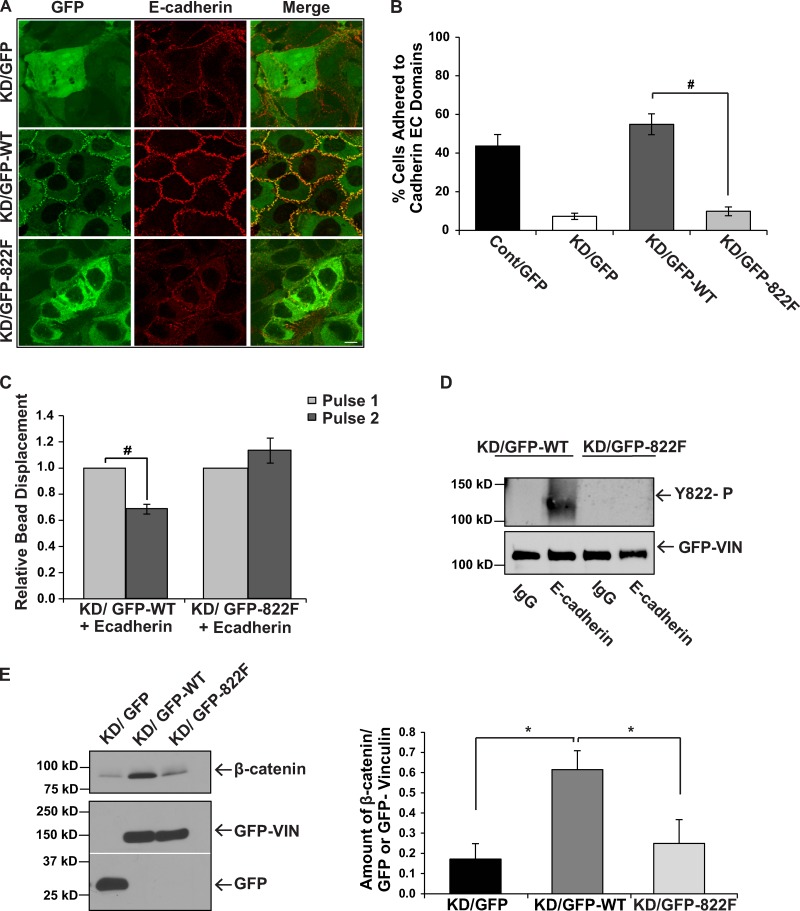
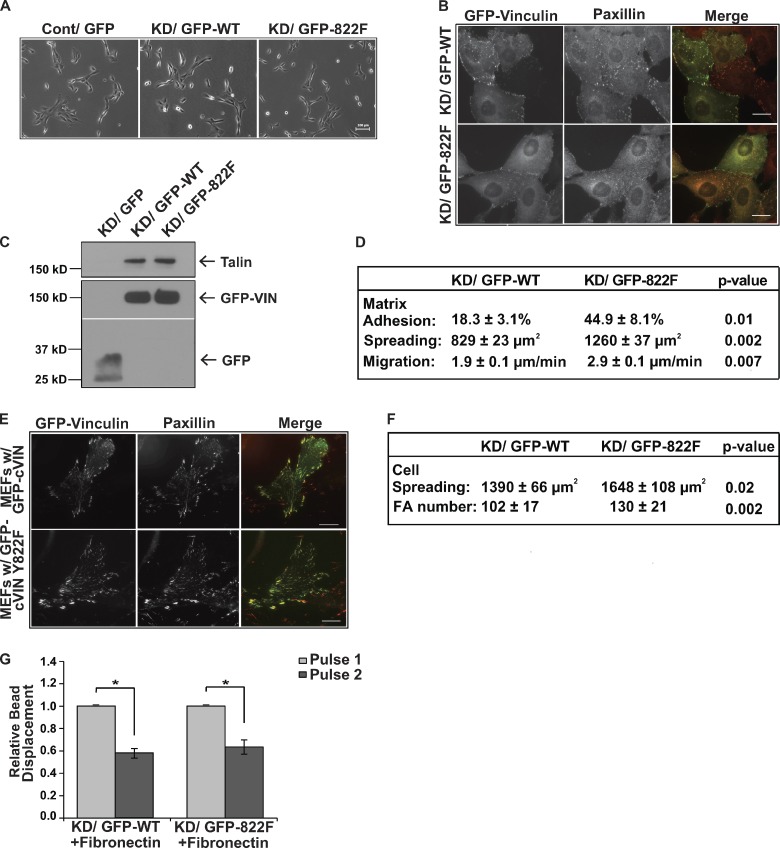
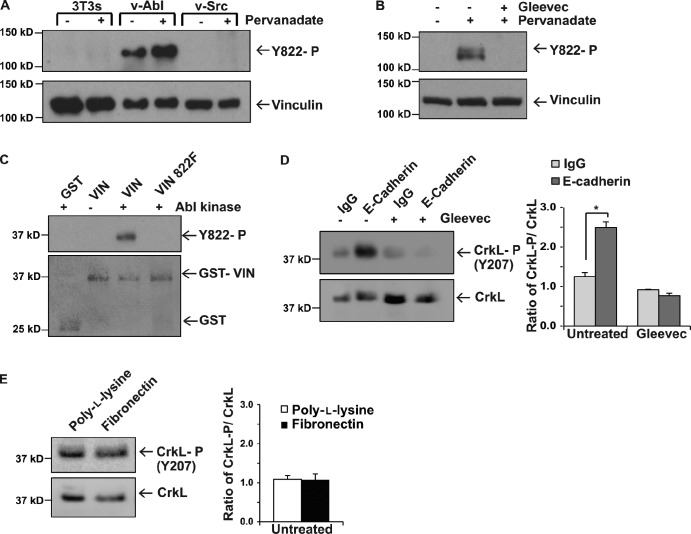
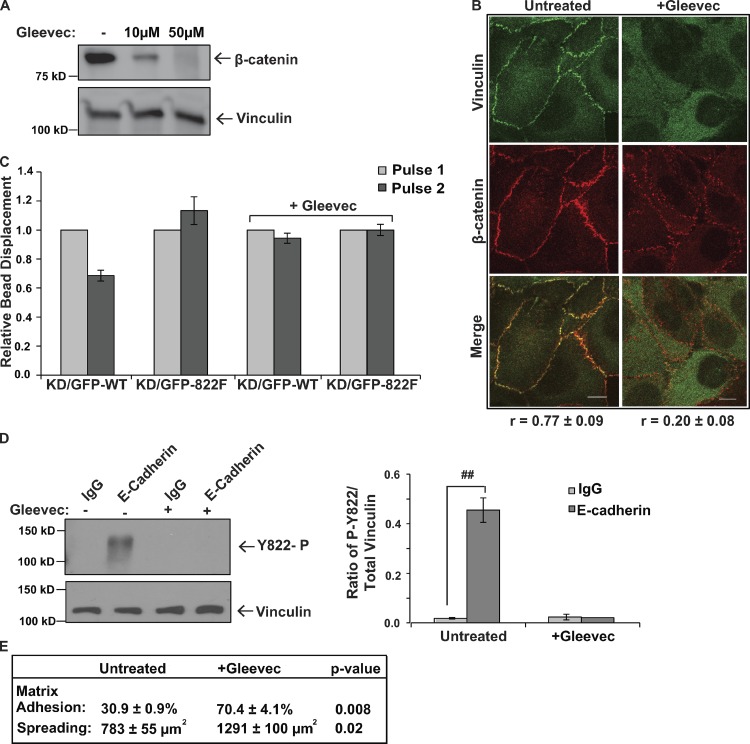
Cells experience mechanical forces throughout their lifetimes. Vinculin is critical for transmitting these forces, yet how it achieves its distinct functions at cell-cell and cell-matrix adhesions remains unanswered. Here, we show vinculin is phosphorylated at Y822 in cell-cell, but not cell-matrix, adhesions. Phosphorylation at Y822 was elevated when forces were applied to E-cadherin and was required for vinculin to integrate into the cadherin complex. The mutation Y822F ablated these activities and prevented cells from stiffening in response to forces on E-cadherin. In contrast, Y822 phosphorylation was not required for vinculin functions in cell-matrix adhesions, including integrin-induced cell stiffening. Finally, forces applied to E-cadherin activated Abelson (Abl) tyrosine kinase to phosphorylate vinculin; Abl inhibition mimicked the loss of vinculin phosphorylation. These data reveal an unexpected regulatory mechanism in which vinculin Y822 phosphorylation determines whether cadherins transmit force and provides a paradigm for how a shared component of adhesions can produce biologically distinct functions.
Figures
References
-
- Borghi N., Sorokina M., Shcherbakova O.G., Weis W.I., Pruitt B.L., Nelson W.J., Dunn A.R. 2012. E-cadherin is under constitutive actomyosin-generated tension that is increased at cell-cell contacts upon externally applied stretch. Proc. Natl. Acad. Sci. USA. 109:12568–12573 10.1073/pnas.1204390109 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
