

Expression of Caveolin 1 is enhanced by DNA demethylation during adipocyte differentiation. status of insulin signaling
- PMID: 24751908
- PMCID: PMC3994010
- DOI: 10.1371/journal.pone.0095100
Expression of Caveolin 1 is enhanced by DNA demethylation during adipocyte differentiation. status of insulin signaling
Abstract
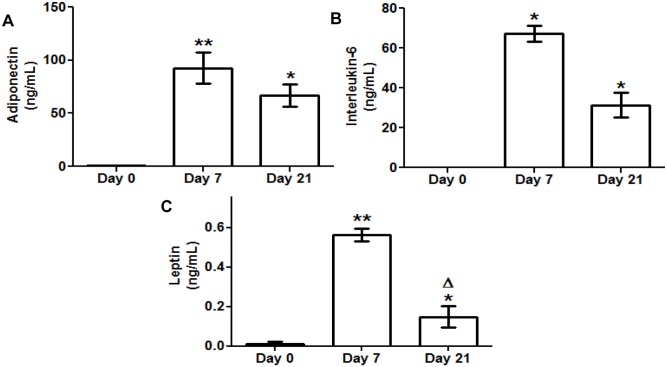
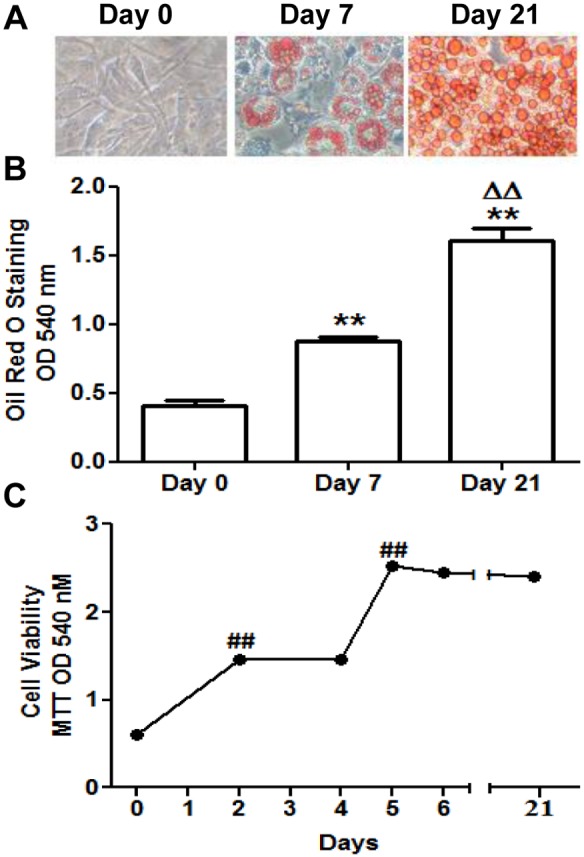
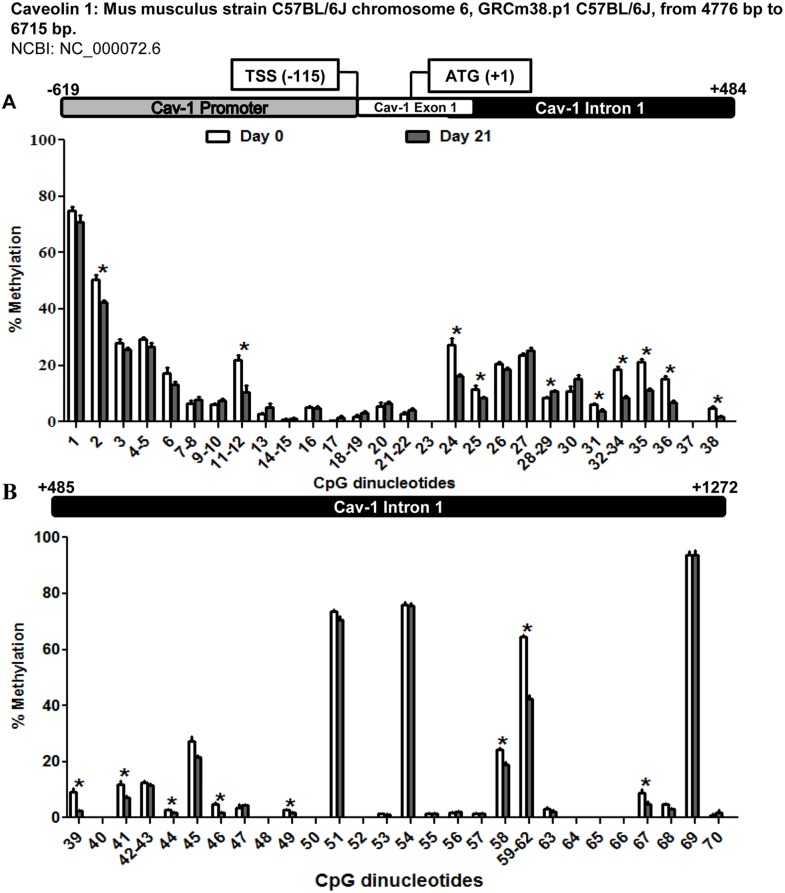
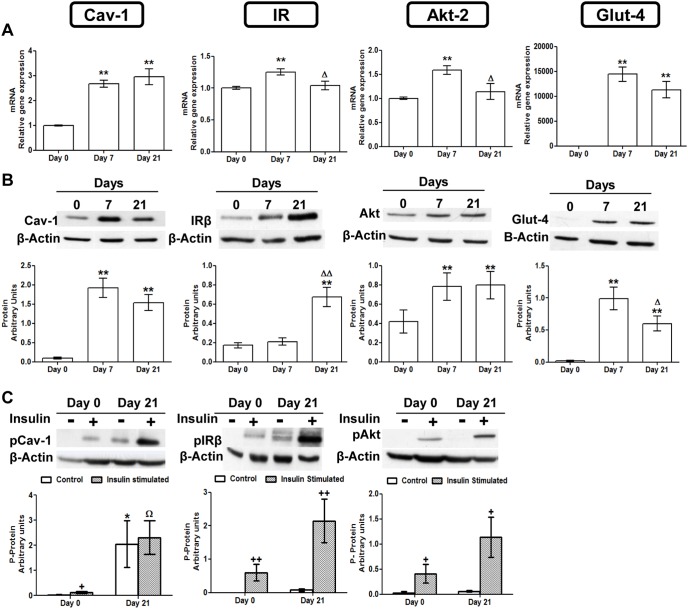
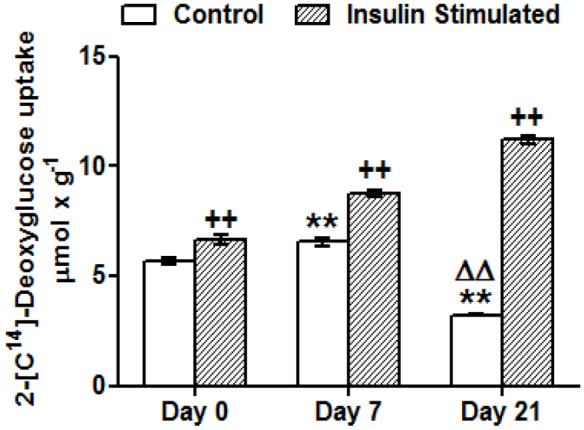
Caveolin 1 (Cav-1) is an essential constituent of adipocyte caveolae which binds the beta subunit of the insulin receptor (IR) and is implicated in the regulation of insulin signaling. We have found that, during adipocyte differentiation of 3T3-L1 cells the promoter, exon 1 and first intron of the Cav-1 gene undergo a demethylation process that is accompanied by a strong induction of Cav-1 expression, indicating that epigenetic mechanisms must have a pivotal role in this differentiation process. Furthermore, IR, PKB-Akt and Glut-4 expression are also increased during the differentiation process suggesting a coordinated regulation with Cav-1. Activation of Cav-1 protein by phosphorylation arises during the differentiation process, yet in fully mature adipocytes insulin is no longer able to significantly increase Cav-1 phosphorylation. However, these long-term differentiated cells are still able to respond adequately to insulin, increasing IR and PKB-Akt phosphorylation and glucose uptake. The activation of Cav-1 during the adipocyte differentiation process could facilitate the maintenance of insulin sensitivity by these fully mature adipocytes isolated from additional external stimuli. However, under the influence of physiological conditions associated to obesity, such as chronic inflammation and hypoxia, insulin sensitivity would finally be compromised.
Conflict of interest statement
Figures
References
-
- Petronis A (2010) Epigenetics as a unifying principle in the aetiology of complex traits and diseases. Nature 465(7299): 721–727. - PubMed
-
- Zhu JG, Ji CB, Zhang CM, Zhu GZ, Shi CM, et al. (2012) Differential DNA methylation status between human preadipocytes and mature adipocytes. Cell Biochem Biophys 63(1): 1–15. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
