

High-affinity glucose transport in Aspergillus nidulans is mediated by the products of two related but differentially expressed genes
- PMID: 24751997
- PMCID: PMC3994029
- DOI: 10.1371/journal.pone.0094662
High-affinity glucose transport in Aspergillus nidulans is mediated by the products of two related but differentially expressed genes
Abstract
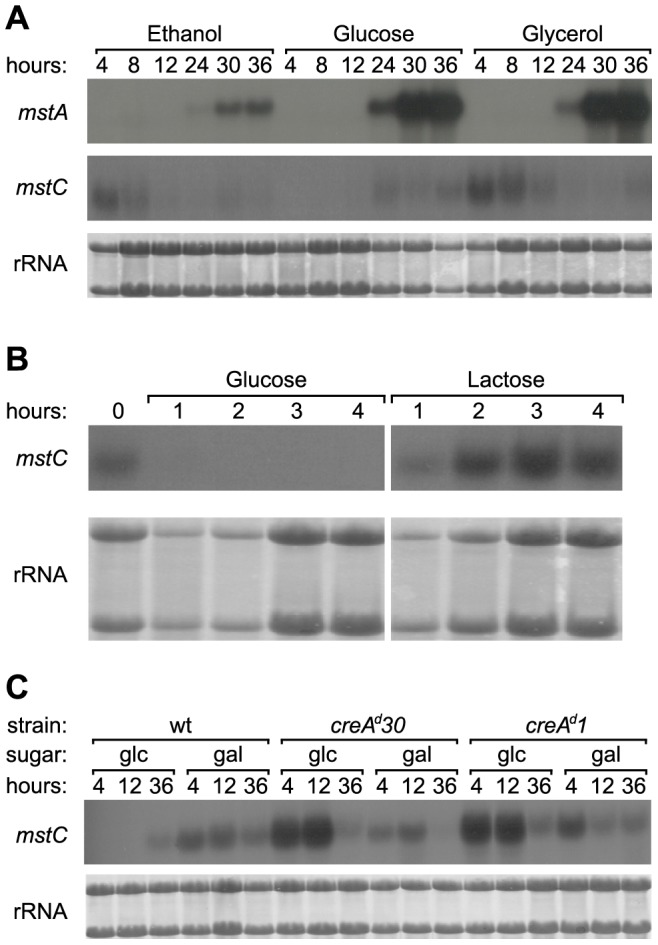
Independent systems of high and low affinity effect glucose uptake in the filamentous fungus Aspergillus nidulans. Low-affinity uptake is known to be mediated by the product of the mstE gene. In the current work two genes, mstA and mstC, have been identified that encode high-affinity glucose transporter proteins. These proteins' primary structures share over 90% similarity, indicating that the corresponding genes share a common origin. Whilst the function of the paralogous proteins is little changed, they differ notably in their patterns of expression. The mstC gene is expressed during the early phases of germination and is subject to CreA-mediated carbon catabolite repression whereas mstA is expressed as a culture tends toward carbon starvation. In addition, various pieces of genetic evidence strongly support allelism of mstC and the previously described locus sorA. Overall, our data define MstC/SorA as a high-affinity glucose transporter expressed in germinating conidia, and MstA as a high-affinity glucose transporter that operates in vegetative hyphae under conditions of carbon limitation.
Conflict of interest statement
Figures





References
-
- Keller NP, Turner G, Bennett JW (2005) Fungal secondary metabolism - from biochemistry to genomics. Nat Rev Microbiol 3: 937–947. - PubMed
-
- Leandro MJ, Fonseca C, Goncalves P (2009) Hexose and pentose transport in ascomycetous yeasts: an overview. FEMS Yeast Res 9: 511–525. - PubMed
-
- Palma M, Goffeau A, Spencer-Martins I, Baret PV (2007) A phylogenetic analysis of the sugar porters in hemiascomycetous yeasts. J Mol Microbiol Biotechnol 12: 241–248. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
