

Genome-wide binding of the CRISPR endonuclease Cas9 in mammalian cells
- PMID: 24752079
- PMCID: PMC4145672
- DOI: 10.1038/nbt.2889
Genome-wide binding of the CRISPR endonuclease Cas9 in mammalian cells
Abstract
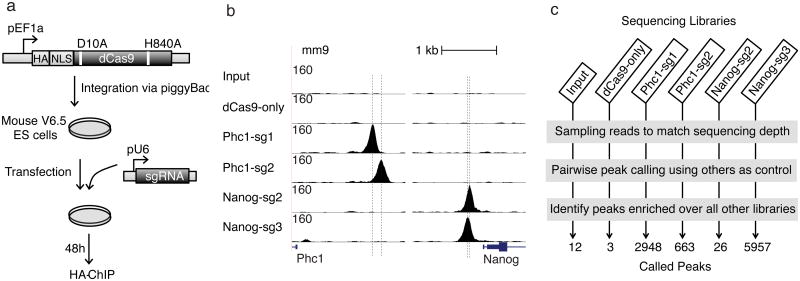
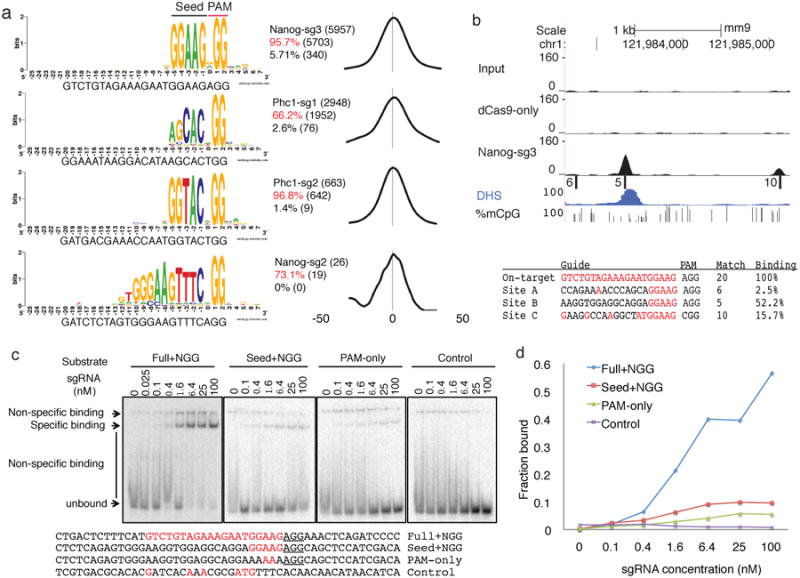
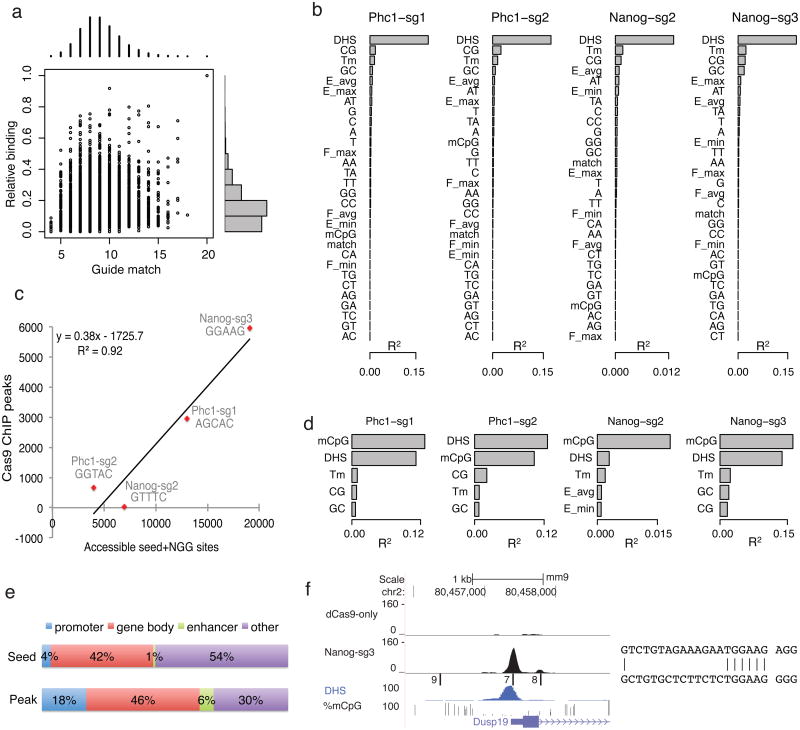
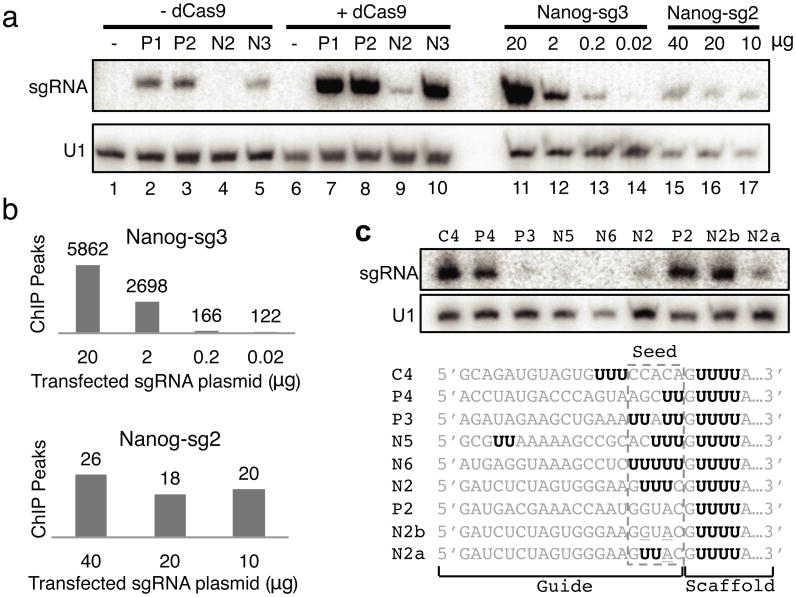
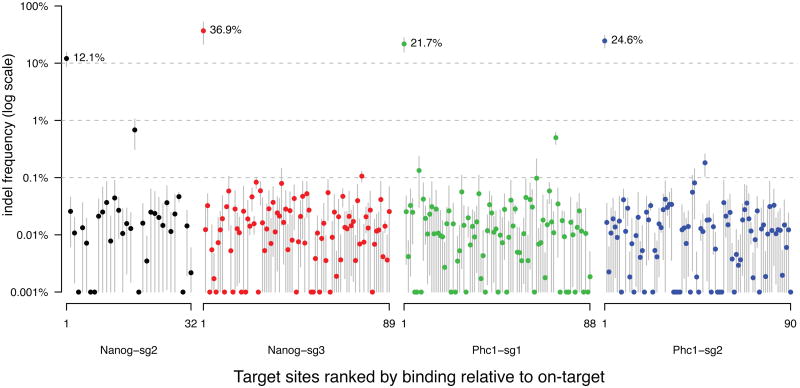
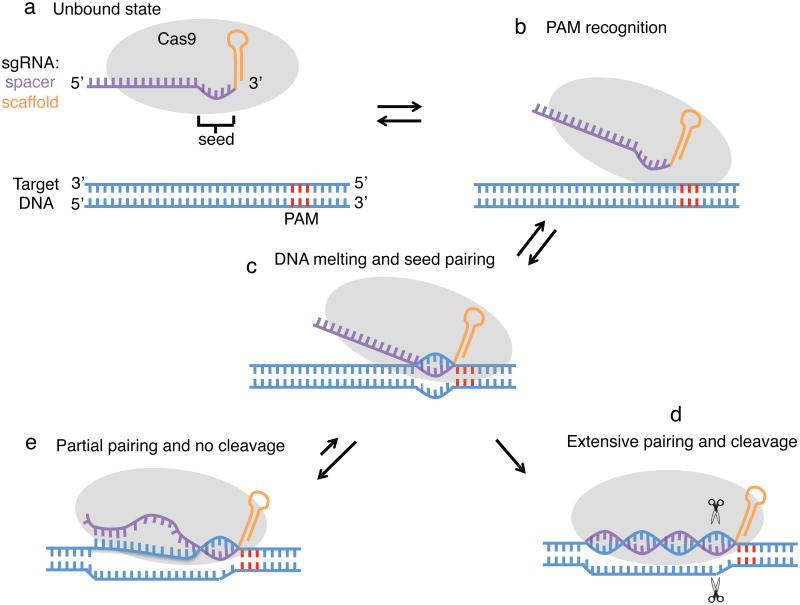
Bacterial type II CRISPR-Cas9 systems have been widely adapted for RNA-guided genome editing and transcription regulation in eukaryotic cells, yet their in vivo target specificity is poorly understood. Here we mapped genome-wide binding sites of a catalytically inactive Cas9 (dCas9) from Streptococcus pyogenes loaded with single guide RNAs (sgRNAs) in mouse embryonic stem cells (mESCs). Each of the four sgRNAs we tested targets dCas9 to between tens and thousands of genomic sites, frequently characterized by a 5-nucleotide seed region in the sgRNA and an NGG protospacer adjacent motif (PAM). Chromatin inaccessibility decreases dCas9 binding to other sites with matching seed sequences; thus 70% of off-target sites are associated with genes. Targeted sequencing of 295 dCas9 binding sites in mESCs transfected with catalytically active Cas9 identified only one site mutated above background levels. We propose a two-state model for Cas9 binding and cleavage, in which a seed match triggers binding but extensive pairing with target DNA is required for cleavage.
Figures
References
-
- Van der Oost J, Jore MM, Westra ER, Lundgren M, Brouns SJJ. CRISPR-based adaptive and heritable immunity in prokaryotes. Trends Biochem Sci. 2009;34:401–7. - PubMed
-
- Deveau H, Garneau JE, Moineau S. CRISPR/Cas system and its role in phage-bacteria interactions. Annu Rev Microbiol. 2010;64:475–93. - PubMed
-
- Horvath P, Barrangou R. CRISPR/Cas, the immune system of bacteria and archaea. Science. 2010;327:167–70. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- SRA/SRP038774
Grants and funding
- T32 GM007287/GM/NIGMS NIH HHS/United States
- R01-CA133404/CA/NCI NIH HHS/United States
- P30 CA014051/CA/NCI NIH HHS/United States
- P30-CA14051/CA/NCI NIH HHS/United States
- R37 HD045022/HD/NICHD NIH HHS/United States
- R01 GM034277/GM/NIGMS NIH HHS/United States
- R01 DK097768/DK/NIDDK NIH HHS/United States
- P01-CA42063/CA/NCI NIH HHS/United States
- 1DP1-MH100706/DP/NCCDPHP CDC HHS/United States
- R01-GM34277/GM/NIGMS NIH HHS/United States
- HHMI/Howard Hughes Medical Institute/United States
- P01 CA042063/CA/NCI NIH HHS/United States
- R37 GM034277/GM/NIGMS NIH HHS/United States
- R01 CA133404/CA/NCI NIH HHS/United States
- DP1 MH100706/MH/NIMH NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
