

Improved HSC reconstitution and protection from inflammatory stress and chemotherapy in mice lacking granzyme B
- PMID: 24752302
- PMCID: PMC4010905
- DOI: 10.1084/jem.20131072
Improved HSC reconstitution and protection from inflammatory stress and chemotherapy in mice lacking granzyme B
Abstract
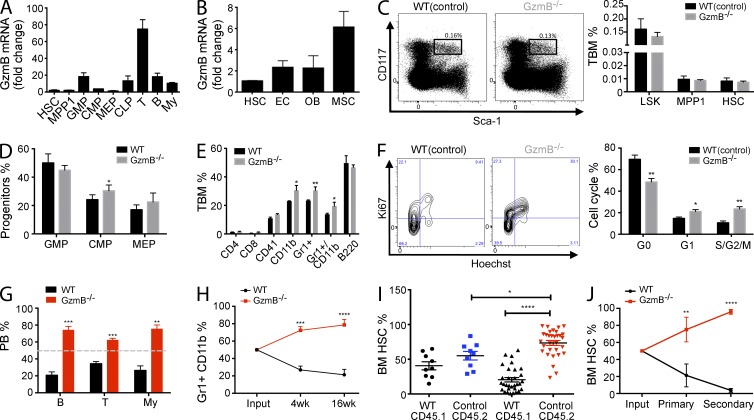
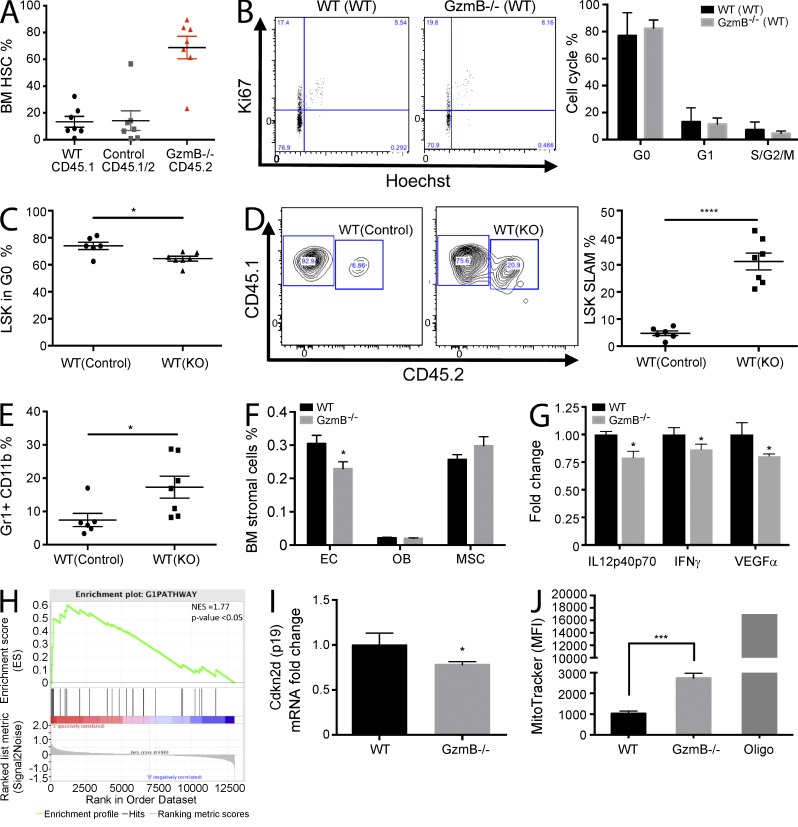
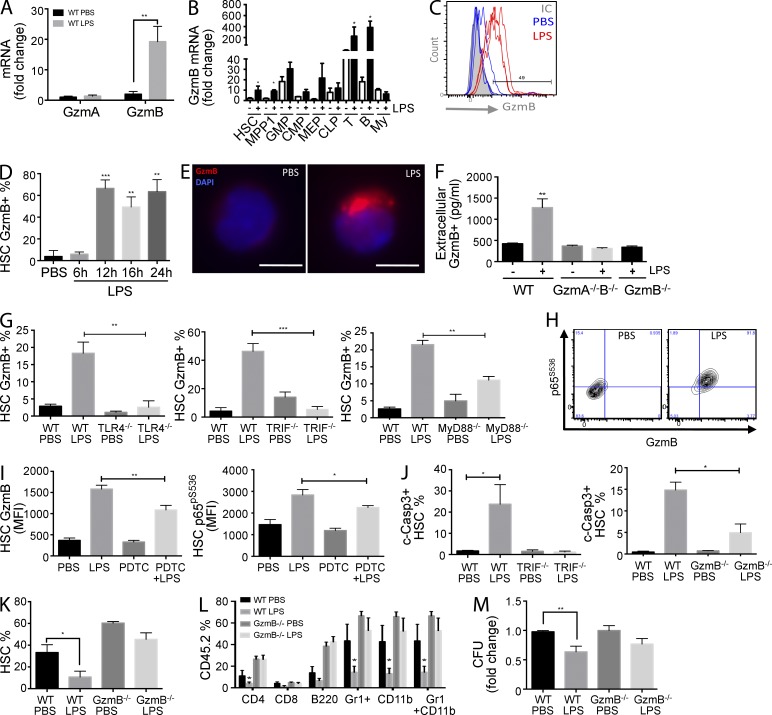
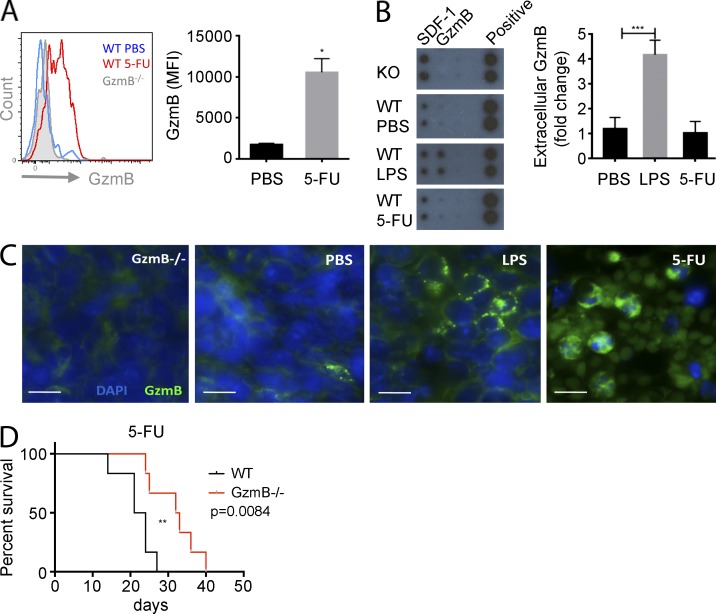
The serine protease granzyme B (GzmB) is stored in the granules of cytotoxic T and NK cells and facilitates immune-mediated destruction of virus-infected cells. In this study, we use genetic tools to report novel roles for GzmB as an important regulator of hematopoietic stem cell (HSC) function in response to stress. HSCs lacking the GzmB gene show improved bone marrow (BM) reconstitution associated with increased HSC proliferation and mitochondrial activity. In addition, recipients deficient in GzmB support superior engraftment of wild-type HSCs compared with hosts with normal BM niches. Stimulation of mice with lipopolysaccharide strongly induced GzmB protein expression in HSCs, which was mediated by the TLR4-TRIF-p65 NF-κB pathway. This is associated with increased cell death and GzmB secretion into the BM environment, suggesting an extracellular role of GzmB in modulating HSC niches. Moreover, treatment with the chemotherapeutic agent 5-fluorouracil (5-FU) also induces GzmB production in HSCs. In this situation GzmB is not secreted, but instead causes cell-autonomous apoptosis. Accordingly, GzmB-deficient mice are more resistant to serial 5-FU treatments. Collectively, these results identify GzmB as a negative regulator of HSC function that is induced by stress and chemotherapy in both HSCs and their niches. Blockade of GzmB production may help to improve hematopoiesis in various situations of BM stress.
Figures
Similar articles
-
Connexin-43 prevents hematopoietic stem cell senescence through transfer of reactive oxygen species to bone marrow stromal cells.Proc Natl Acad Sci U S A. 2012 Jun 5;109(23):9071-6. doi: 10.1073/pnas.1120358109. Epub 2012 May 18. Proc Natl Acad Sci U S A. 2012. PMID: 22611193 Free PMC article.
-
Total body irradiation causes residual bone marrow injury by induction of persistent oxidative stress in murine hematopoietic stem cells.Free Radic Biol Med. 2010 Jan 15;48(2):348-56. doi: 10.1016/j.freeradbiomed.2009.11.005. Epub 2009 Dec 2. Free Radic Biol Med. 2010. PMID: 19925862 Free PMC article.
-
Bone marrow connexin-43 expression is critical for hematopoietic regeneration after chemotherapy.Cell Commun Adhes. 2005 Jul-Dec;12(5-6):307-17. doi: 10.1080/15419060500514200. Cell Commun Adhes. 2005. PMID: 16531325
-
Extravascular coagulation in hematopoietic stem and progenitor cell regulation.Blood. 2018 Jul 12;132(2):123-131. doi: 10.1182/blood-2017-12-768986. Epub 2018 Jun 4. Blood. 2018. PMID: 29866813 Free PMC article. Review.
-
The biological function of Serpinb9 and Serpinb9-based therapy.Front Immunol. 2024 Jun 20;15:1422113. doi: 10.3389/fimmu.2024.1422113. eCollection 2024. Front Immunol. 2024. PMID: 38966643 Free PMC article. Review.
Cited by
-
Constitutive Activation of NIK Impairs the Self-Renewal of Hematopoietic Stem/Progenitor Cells and Induces Bone Marrow Failure.Stem Cells. 2017 Mar;35(3):777-786. doi: 10.1002/stem.2523. Epub 2016 Nov 2. Stem Cells. 2017. PMID: 27733012 Free PMC article.
-
Cutting the brakes on hematopoietic regeneration by blocking TGFβ to limit chemotherapy-induced myelosuppression.Mol Cell Oncol. 2015 Apr 18;2(3):e978703. doi: 10.4161/23723556.2014.978703. eCollection 2015 Jul-Sep. Mol Cell Oncol. 2015. PMID: 27308454 Free PMC article. Review.
-
Yap1-Scribble polarization is required for hematopoietic stem cell division and fate.Blood. 2020 Oct 15;136(16):1824-1836. doi: 10.1182/blood.2019004113. Blood. 2020. PMID: 32483624 Free PMC article.
-
Competitive sgRNA Screen Identifies p38 MAPK as a Druggable Target to Improve HSPC Engraftment.Cells. 2020 Sep 29;9(10):2194. doi: 10.3390/cells9102194. Cells. 2020. PMID: 33003308 Free PMC article.
-
Regulation of stress-induced hematopoiesis.Curr Opin Hematol. 2015 Jul;22(4):286-92. doi: 10.1097/MOH.0000000000000149. Curr Opin Hematol. 2015. PMID: 26049748 Free PMC article. Review.
References
-
- Birdsall H.H., Porter W.J., Green D.M., Rubio J., Trial J., Rossen R.D. 2004. Impact of fibronectin fragments on the transendothelial migration of HIV-infected leukocytes and the development of subendothelial foci of infectious leukocytes. J. Immunol. 173:2746–2754 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
