

The canonical wnt signal restricts the glycogen synthase kinase 3/fbw7-dependent ubiquitination and degradation of eya1 phosphatase
- PMID: 24752894
- PMCID: PMC4054306
- DOI: 10.1128/MCB.00104-14
The canonical wnt signal restricts the glycogen synthase kinase 3/fbw7-dependent ubiquitination and degradation of eya1 phosphatase
Abstract
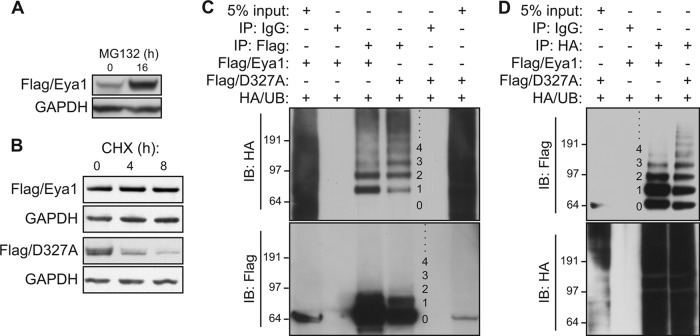
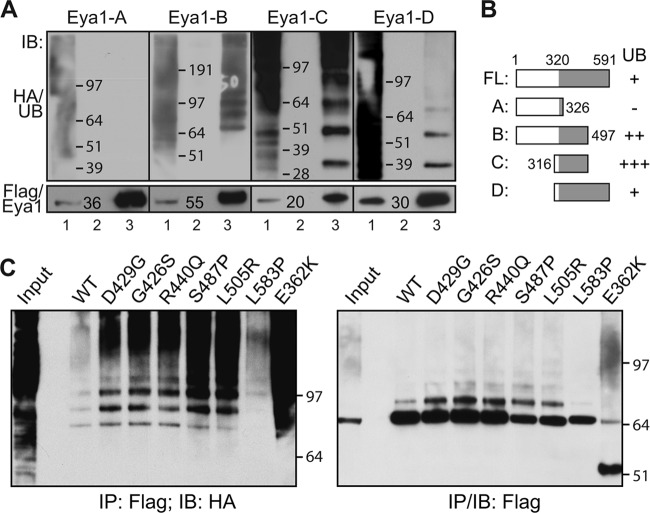
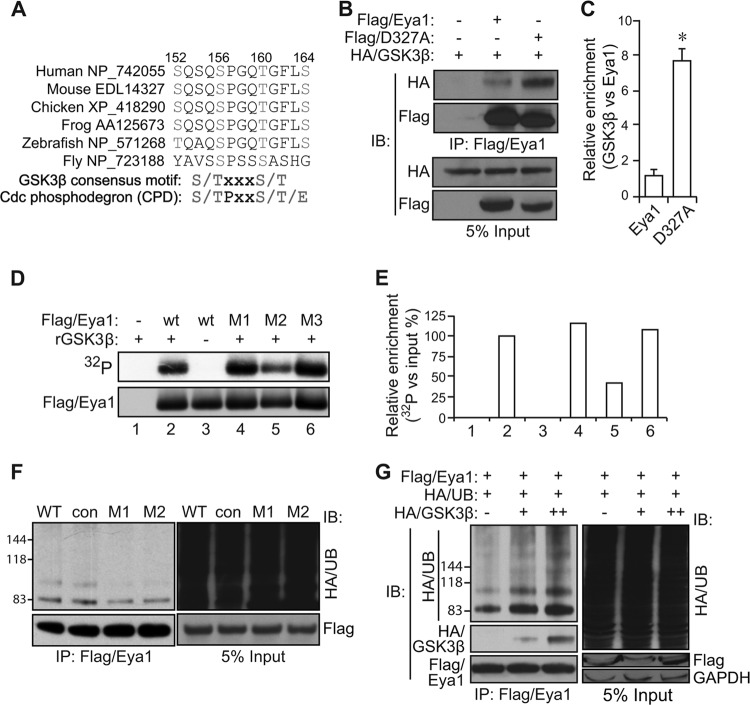
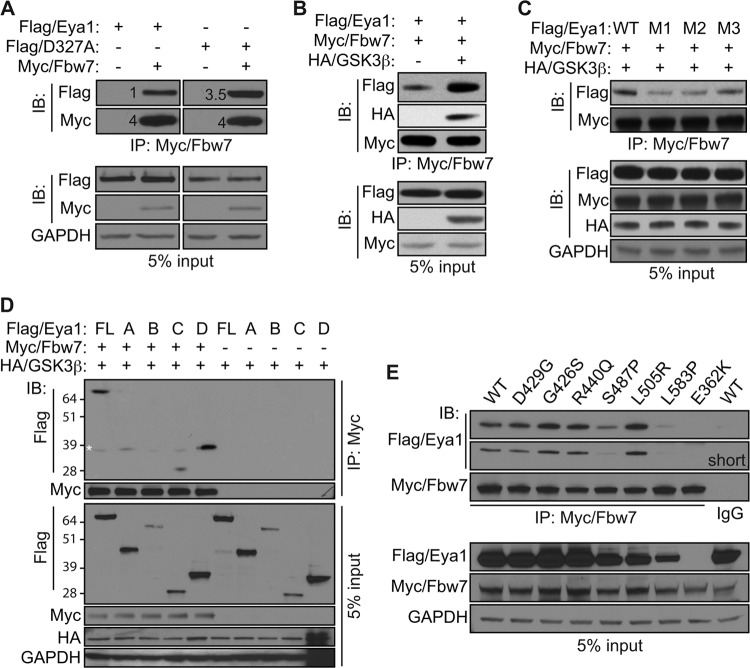
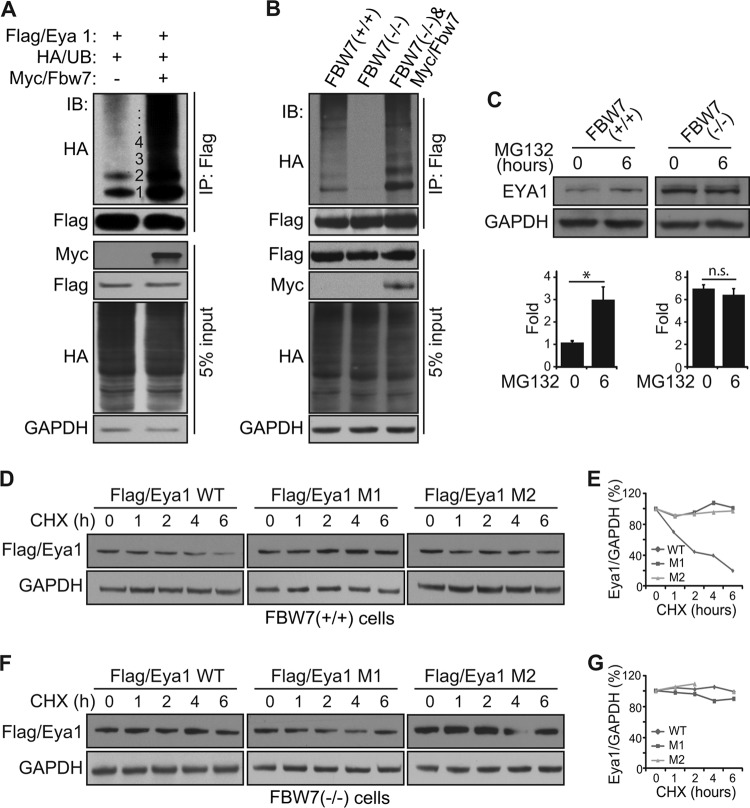
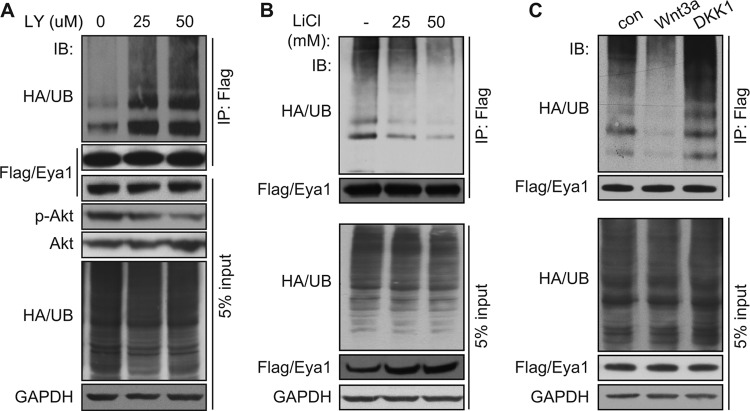
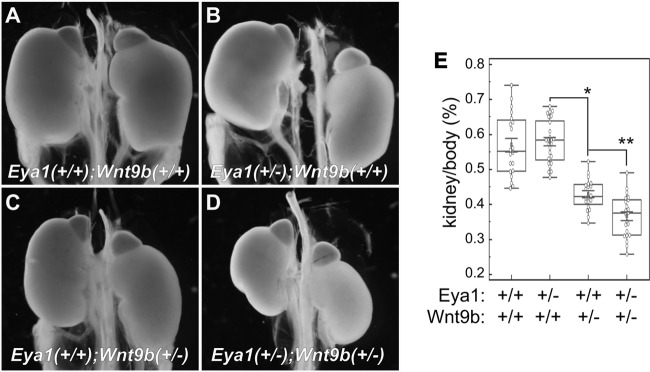
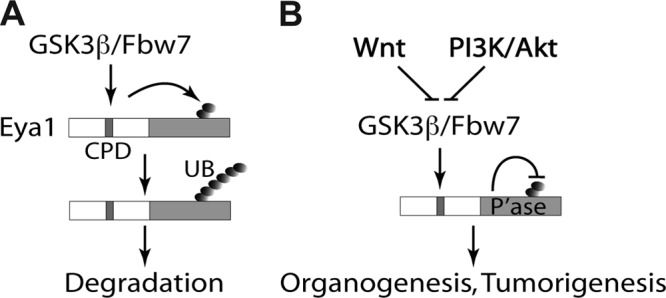
Haploinsufficiency of Eya1 causes the branchio-oto-renal (BOR) syndrome, and abnormally high levels of Eya1 are linked to breast cancer progression and poor prognosis. Therefore, regulation of Eya1 activity is key to its tissue-specific functions and oncogenic activities. Here, we show that Eya1 is posttranslationally modified by ubiquitin and that its ubiquitination level is self-limited to prevent premature degradation. Eya1 has an evolutionarily conserved CDC4 phosphodegron (CPD) signal, a target site of glycogen synthase kinase 3 (GSK3) kinase and Fbw7 ubiquitin ligase, which is required for Eya1 ubiquitination. Genetic deletion of Fbw7 and pharmacological inhibition of GSK3 significantly decrease Eya1 ubiquitination. Conversely, activation of the phosphatidylinositol 3-kinase (PI3K)/Akt and the canonical Wnt signal suppresses Eya1 ubiquitination. Compound Eya1(+/-); Wnt9b(+/-) mutants exhibit an increased penetrance of renal defect, indicating that they function in the same genetic pathway in vivo. Together, these findings reveal that the canonical Wnt and PI3K/Akt signal pathways restrain the GSK3/Fbw7-dependent Eya1 ubiquitination, and they further suggest that dysregulation of this novel axis contributes to tumorigenesis.
Copyright © 2014, American Society for Microbiology. All Rights Reserved.
Figures
References
-
- Bonini NM, Bui QT, Gray-Board GL, Warrick JM. 1997. The Drosophila eyes absent gene directs ectopic eye formation in a pathway conserved between flies and vertebrates. Development 124:4819–4826 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials