

An N-terminal threonine mutation produces an efflux-favorable, sodium-primed conformation of the human dopamine transporter
- PMID: 24753048
- PMCID: PMC4053999
- DOI: 10.1124/mol.114.091926
An N-terminal threonine mutation produces an efflux-favorable, sodium-primed conformation of the human dopamine transporter
Abstract
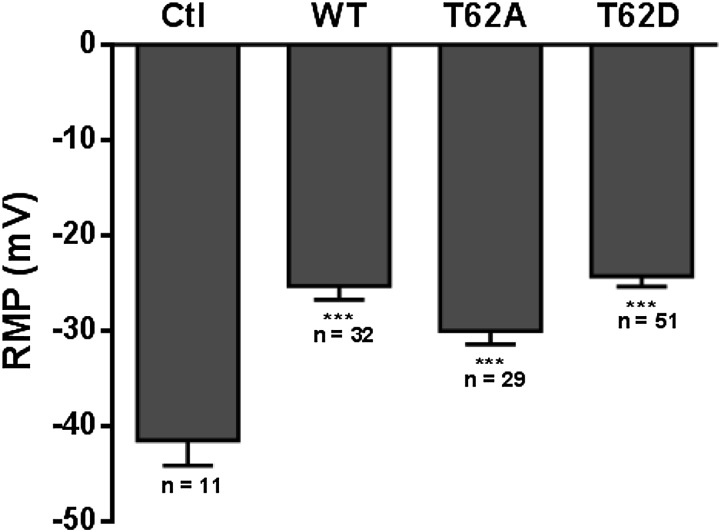
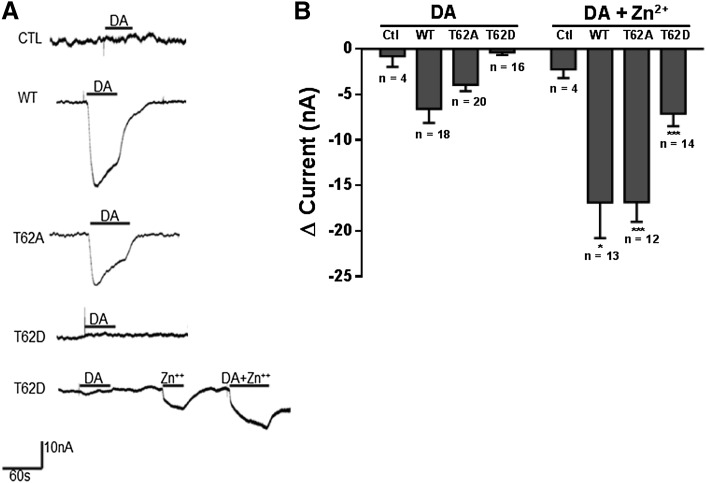
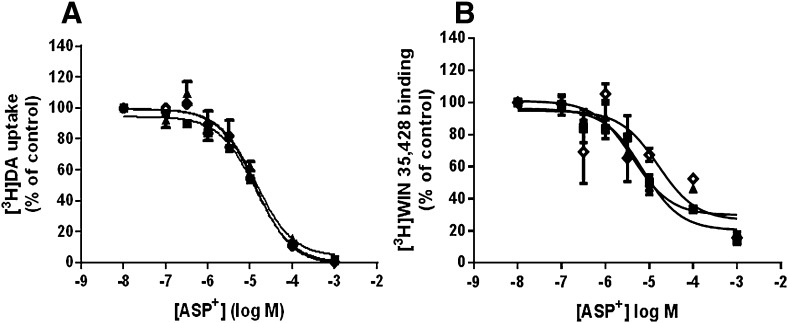
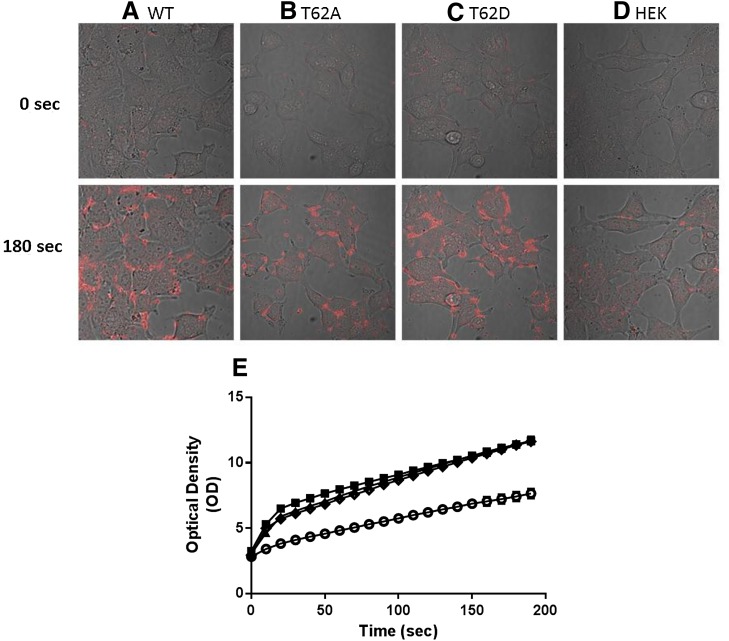
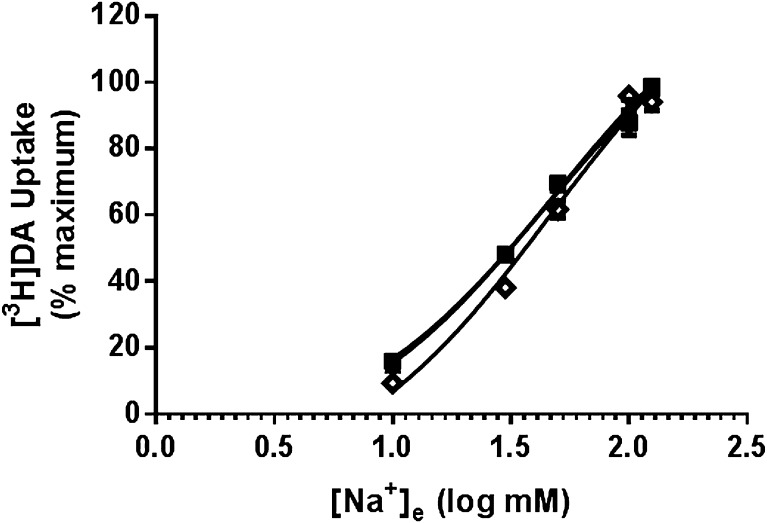
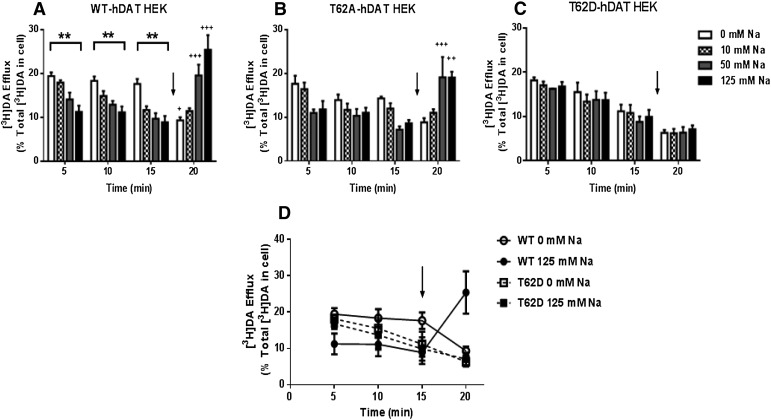
The dopamine transporter (DAT) reversibly transports dopamine (DA) through a series of conformational transitions. Alanine (T62A) or aspartate (T62D) mutagenesis of Thr62 revealed T62D-human (h)DAT partitions in a predominately efflux-preferring conformation. Compared with wild-type (WT), T62D-hDAT exhibits reduced [(3)H]DA uptake and enhanced baseline DA efflux, whereas T62A-hDAT and WT-hDAT function in an influx-preferring conformation. We now interrogate the basis of the mutants' altered function with respect to membrane conductance and Na(+) sensitivity. The hDAT constructs were expressed in Xenopus oocytes to investigate if heightened membrane potential would explain the efflux characteristics of T62D-hDAT. In the absence of substrate, all constructs displayed identical resting membrane potentials. Substrate-induced inward currents were present in oocytes expressing WT- and T62A-hDAT but not T62D-hDAT, suggesting equal bidirectional ion flow through T62D-hDAT. Utilization of the fluorescent DAT substrate ASP(+) [4-(4-(dimethylamino)styryl)-N-methylpyridinium] revealed that T62D-hDAT accumulates substrate in human embryonic kidney (HEK)-293 cells when the substrate is not subject to efflux. Extracellular sodium (Na(+) e) replacement was used to evaluate sodium gradient requirements for DAT transport functions. The EC50 for Na(+) e stimulation of [(3)H]DA uptake was identical in all constructs expressed in HEK-293 cells. As expected, decreasing [Na(+)]e stimulated [(3)H]DA efflux in WT- and T62A-hDAT cells. Conversely, the elevated [(3)H]DA efflux in T62D-hDAT cells was independent of Na(+) e and commensurate with [(3)H]DA efflux attained in WT-hDAT cells, either by removal of Na(+) e or by application of amphetamine. We conclude that T62D-hDAT represents an efflux-willing, Na(+)-primed orientation-possibly representing an experimental model of the conformational impact of amphetamine exposure to hDAT.
Copyright © 2014 by The American Society for Pharmacology and Experimental Therapeutics.
Figures

Similar articles
-
A juxtamembrane mutation in the N terminus of the dopamine transporter induces preference for an inward-facing conformation.Mol Pharmacol. 2009 Mar;75(3):514-24. doi: 10.1124/mol.108.048744. Epub 2008 Dec 19. Mol Pharmacol. 2009. PMID: 19098122 Free PMC article.
-
Site-directed mutations near transmembrane domain 1 alter conformation and function of norepinephrine and dopamine transporters.Mol Pharmacol. 2011 Mar;79(3):520-32. doi: 10.1124/mol.110.069039. Epub 2010 Dec 13. Mol Pharmacol. 2011. PMID: 21149640 Free PMC article.
-
Amphetamine regulation of dopamine transport. Combined measurements of transporter currents and transporter imaging support the endocytosis of an active carrier.J Biol Chem. 2004 Mar 5;279(10):8966-75. doi: 10.1074/jbc.M303976200. Epub 2003 Dec 29. J Biol Chem. 2004. PMID: 14699142
-
Anomalous dopamine release associated with a human dopamine transporter coding variant.J Neurosci. 2008 Jul 9;28(28):7040-6. doi: 10.1523/JNEUROSCI.0473-08.2008. J Neurosci. 2008. PMID: 18614672 Free PMC article.
-
Impact of disruption of secondary binding site S2 on dopamine transporter function.J Neurochem. 2016 Sep;138(5):694-9. doi: 10.1111/jnc.13704. Epub 2016 Jul 14. J Neurochem. 2016. PMID: 27315582 Free PMC article. Review.
Cited by
-
Studies of the biogenic amine transporters 15. Identification of novel allosteric dopamine transporter ligands with nanomolar potency.J Pharmacol Exp Ther. 2015 Jun;353(3):529-38. doi: 10.1124/jpet.114.222299. Epub 2015 Mar 18. J Pharmacol Exp Ther. 2015. PMID: 25788711 Free PMC article.
-
Kinase-dependent Regulation of Monoamine Neurotransmitter Transporters.Pharmacol Rev. 2016 Oct;68(4):888-953. doi: 10.1124/pr.115.012260. Pharmacol Rev. 2016. PMID: 27591044 Free PMC article. Review.
-
Chloride requirement for monoamine transporters.Pflugers Arch. 2016 Mar;468(3):503-11. doi: 10.1007/s00424-015-1783-4. Epub 2016 Jan 22. Pflugers Arch. 2016. PMID: 26794730 Free PMC article. Review.
-
Dopamine Transporter Activity Is Modulated by α-Synuclein.J Biol Chem. 2015 Dec 4;290(49):29542-54. doi: 10.1074/jbc.M115.691592. Epub 2015 Oct 6. J Biol Chem. 2015. PMID: 26442590 Free PMC article.
-
The dopamine transporter: An unrecognized nexus for dysfunctional peripheral immunity and signaling in Parkinson's Disease.Brain Behav Immun. 2018 May;70:21-35. doi: 10.1016/j.bbi.2018.03.020. Epub 2018 Mar 15. Brain Behav Immun. 2018. PMID: 29551693 Free PMC article. Review.
References
-
- Chen N, Rickey J, Berfield JL, Reith ME. (2004a) Aspartate 345 of the dopamine transporter is critical for conformational changes in substrate translocation and cocaine binding. J Biol Chem 279:5508–5519 - PubMed
-
- Chen, NH, Reith, ME, and Quick, MW (2004b). Synaptic uptake and beyond: the sodium- and chloride-dependent neurotransmitter transporter family SLC6. Pflugers Arch. 447:519–531. - PubMed
-
- DeFelice LJ, Goswami T. (2007) Transporters as channels. Annu Rev Physiol 69:87–112 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
