

Prelamin A accumulation and stress conditions induce impaired Oct-1 activity and autophagy in prematurely aged human mesenchymal stem cell
- PMID: 24753226
- PMCID: PMC4032794
- DOI: 10.18632/aging.100651
Prelamin A accumulation and stress conditions induce impaired Oct-1 activity and autophagy in prematurely aged human mesenchymal stem cell
Abstract
Aging, a time-dependent functional decline of biological processes, is the primary risk factor in developing diseases such as cancer, cardiovascular or degenerative diseases. There is a real need to understand the human aging process in order to increase the length of disease-free life, also known as "health span". Accumulation of progerin and prelamin A are the hallmark of a group of premature aging diseases but have also been found during normal cellular aging strongly suggesting similar mechanisms between healthy aging and LMNA-linked progeroid syndromes. How this toxic accumulation contributes to aging (physiological or pathological) remains unclear. Since affected tissues in age-associated disorders and in pathological aging are mainly of mesenchymal origin we propose a model of human aging based on mesenchymal stem cells (hMSCs) which accumulate prelamin A. We demonstrate that prelamin A-accumulating hMSCs have a premature aging phenotype which affects their functional competence in vivo. The combination of prelamin A accumulation and stress conditions enhance the aging phenotype by dysregulating the activity of the octamer binding protein Oct-1This experimental model has been fundamental to identify a new role for Oct-1 in hMSCs aging.
Conflict of interest statement
No conflict of interest could be disclosed for any author.
Figures

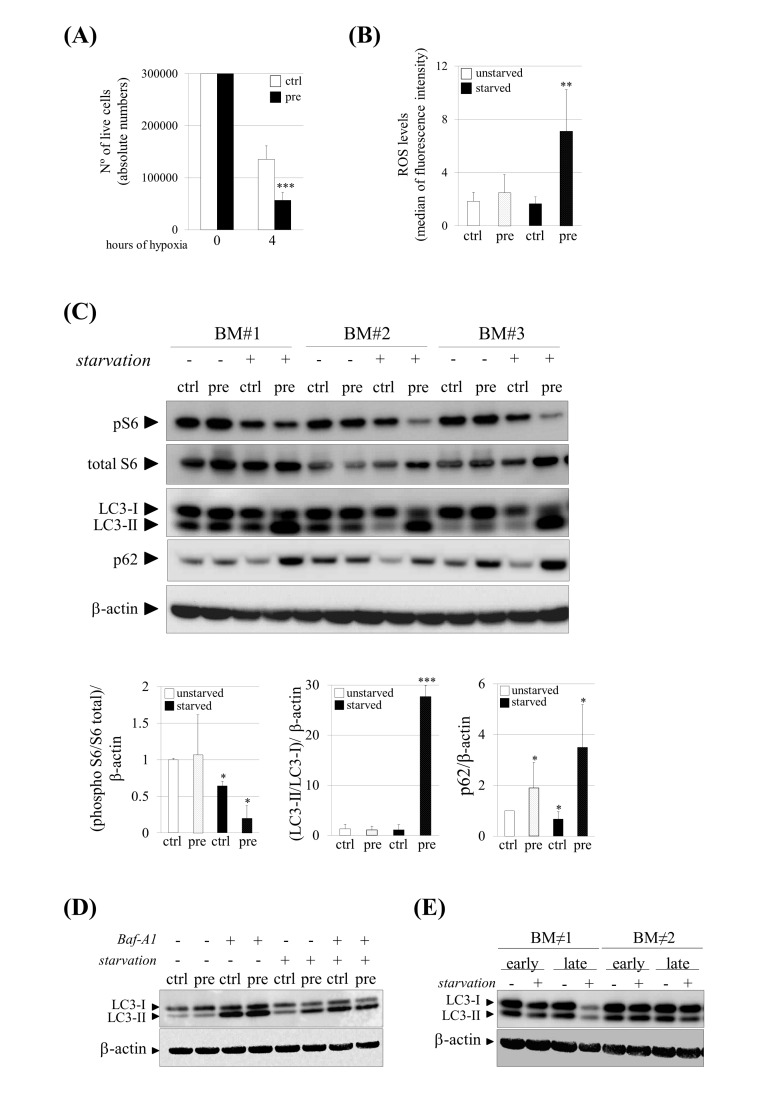


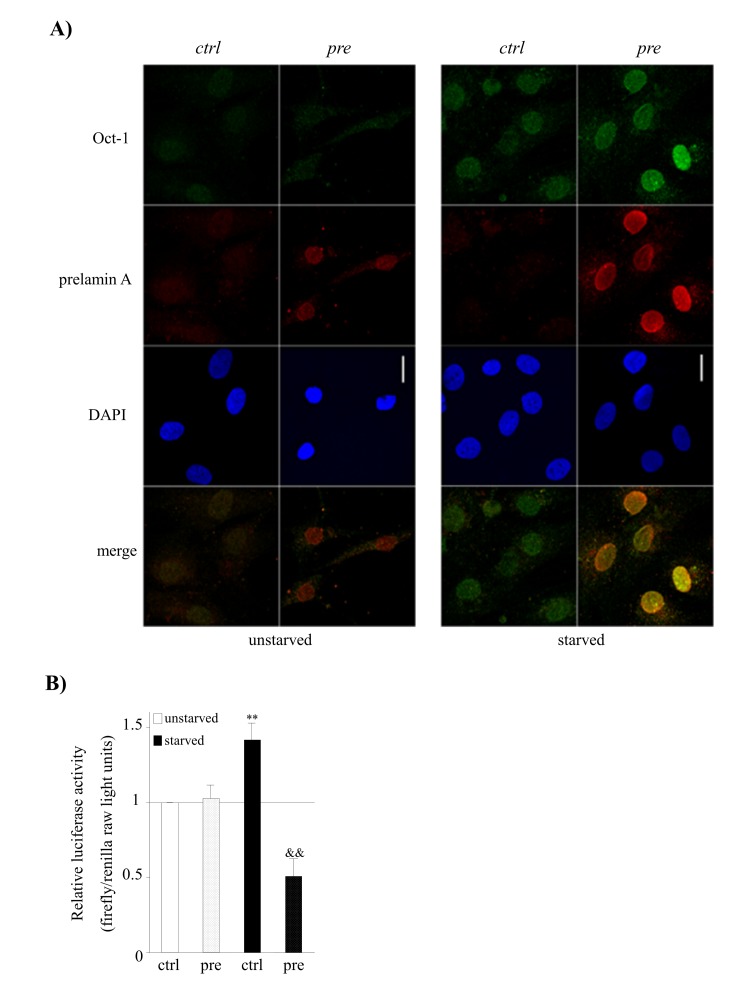

References
-
- Ragnauth CD, Warren DT, Liu Y, McNair R, Tajsic T, Figg N, Shroff R, Skepper J, Shanahan CM. Prelamin A acts to accelerate smooth muscle cell senescence and is a novel biomarker of human vascular aging. Circulation. 2010;121:2200–2210. - PubMed
-
- De Sandre-Giovannoli A, Bernard R, Cau P, Navarro C, Amiel J, Boccaccio I, Lyonnet S, Stewart CL, Munnich A, Le Merrer M, Lévy N. Lamin a truncation in Hutchinson-Gilford progeria. Science. 2003;300:2055. - PubMed
-
- Capanni C, Mattioli E, Columbaro M, Lucarelli E, Parnaik VK, Novelli G, Wehnert M, Cenni V, Maraldi NM, Squarzoni S, Lattanzi G. Altered pre-lamin A processing is a common mechanism leading to lipodystrophy. Hum Mol Genet. 2005;14:1489–1502. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
