

miR-200c regulates IL8 expression by targeting IKBKB: a potential mediator of inflammation in leiomyoma pathogenesis
- PMID: 24755559
- PMCID: PMC3995706
- DOI: 10.1371/journal.pone.0095370
miR-200c regulates IL8 expression by targeting IKBKB: a potential mediator of inflammation in leiomyoma pathogenesis
Abstract
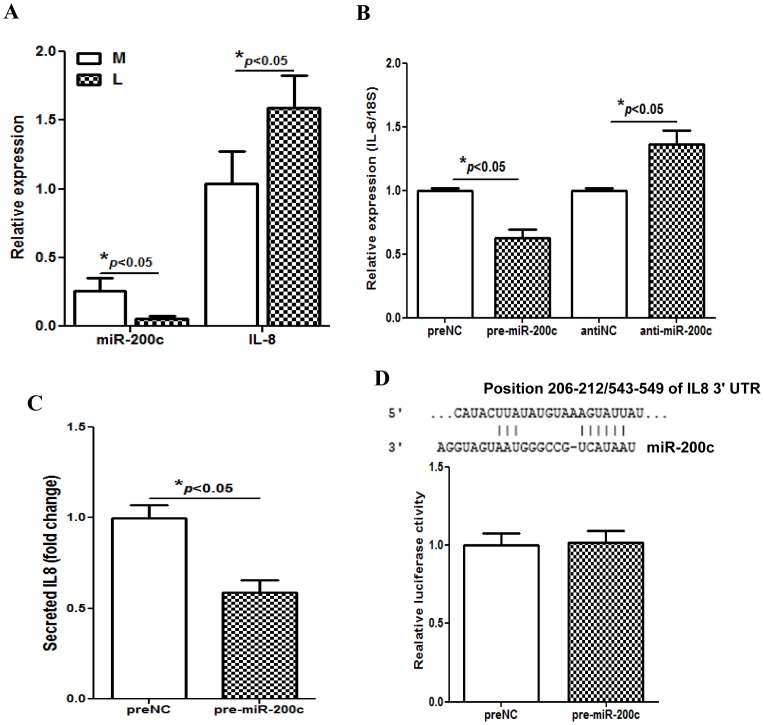
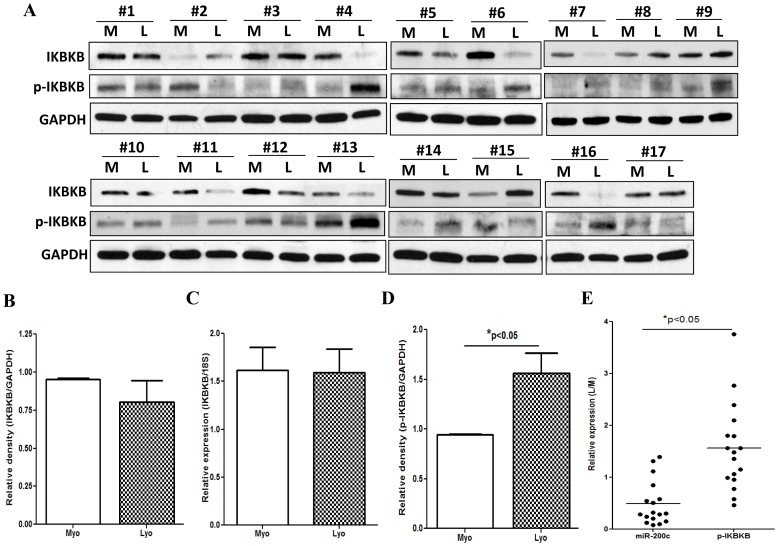
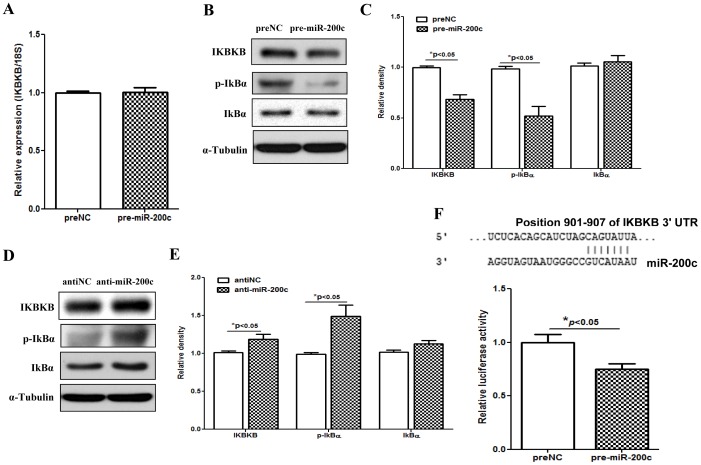
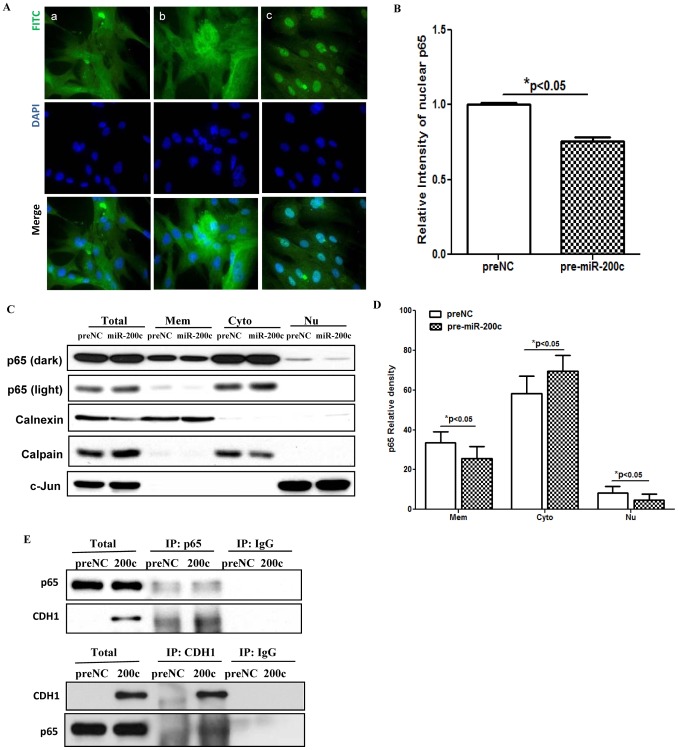
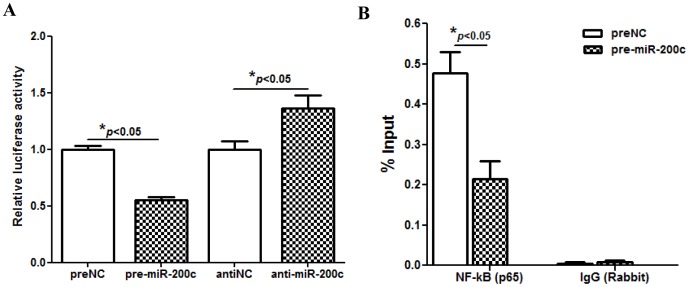
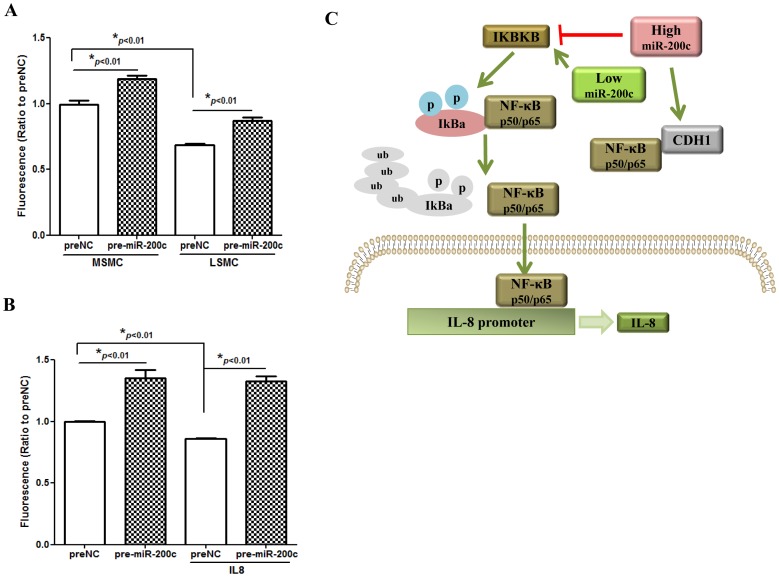
We have previously reported that leiomyoma expressed lower levels of miR-200c and elevated IL8 as compared to paired myometrium. Here we addressed the regulatory functions of miR-200c on the expression of inflammatory mediators and cellular viability using leiomyomas and paired myometrium and their isolated primary smooth muscle cells. Our results indicated that gain-of function or knockdown of miR-200c in leiomyoma smooth muscle cells (LSMC) regulated IL8 mRNA and protein expression through direct targeting of IKBKB and alteration of NF-kB activity. Additionally, leiomyoma expressed higher levels of phosphorylated IKBKB with no significant difference in the level of IKBKB mRNA and protein as compared to matched myometrium. Gain-of function of miR-200c in LSMC resulted in decreased IkBα phosphorylation and p65 nuclear translocation, which led to decreased p65 transcriptional activity of IL8 promoter, and increased caspase 3/7 activity which was not reversible following IL8 restoration. Collectively, our results suggest that NF-κB signaling pathway is a target of miR-200c regulatory function, and low level of miR-200c expression in leiomyoma by transcriptional regulation of inflammatory mediators such as IL8, in part account for development of leiomyomas.
Conflict of interest statement
Figures
References
-
- Schafer M, Werner S (2008) Cancer as an overhealing wound: an old hypothesis revisited. Nat Rev Mol Cell Biol 9: 628–638. - PubMed
-
- Fabian MR, Sonenberg N, Filipowicz W (2010) Regulation of mRNA translation and stability by microRNAs. Annu Rev Biochem 79: 351–379. - PubMed
-
- Huntzinger E, Izaurralde E (2011) Gene silencing by microRNAs: contributions of translational repression and mRNA decay. Nat Rev Genet 12: 99–110. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
