

A method to assess target gene involvement in angiogenesis in vitro and in vivo using lentiviral vectors expressing shRNA
- PMID: 24759702
- PMCID: PMC3997504
- DOI: 10.1371/journal.pone.0096036
A method to assess target gene involvement in angiogenesis in vitro and in vivo using lentiviral vectors expressing shRNA
Abstract
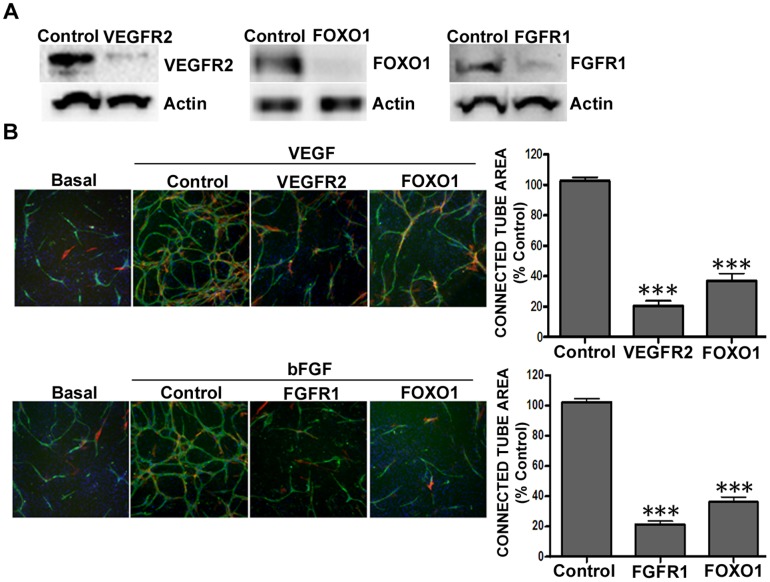
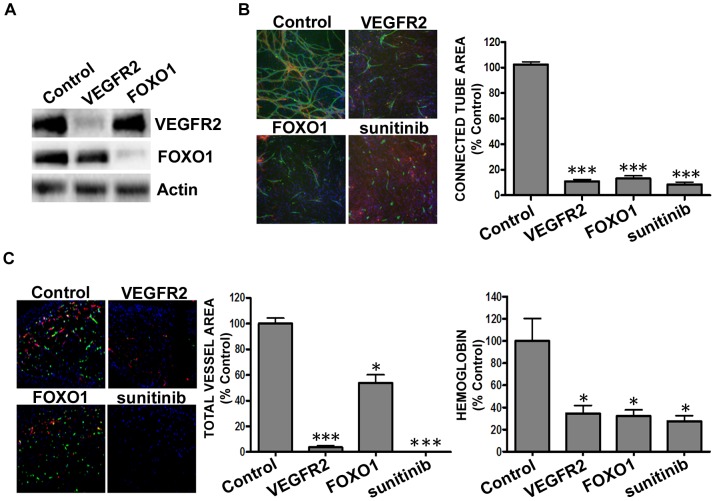
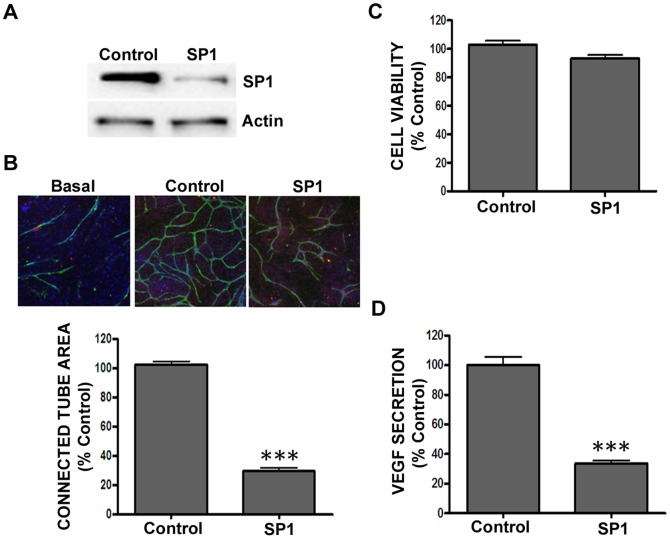
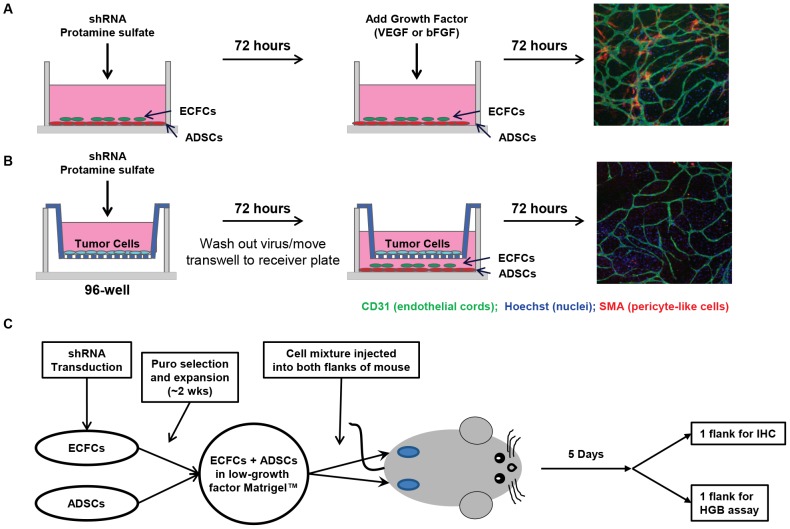
Current methods to study angiogenesis in cancer growth and development can be difficult and costly, requiring extensive use of in vivo methodologies. Here, we utilized an in vitro adipocyte derived stem cell and endothelial colony forming cell (ADSC/ECFC) co-culture system to investigate the effect of lentiviral-driven shRNA knockdown of target genes compared to a non-targeting shRNA control on cord formation using High Content Imaging. Cord formation was significantly reduced following knockdown of the VEGF receptor VEGFR2 in VEGF-driven cord formation and the FGF receptor FGFR1 in basic FGF (bFGF)-driven cord formation. In addition, cord formation was significantly reduced following knockdown of the transcription factor forkhead box protein O1 (FOXO1), a protein with known positive effects on angiogenesis and blood vessel stabilization in VEGF- and bFGF-driven cord formation. Lentiviral shRNA also demonstrated utility for stable knockdown of VEGFR2 and FOXO1 in ECFCs, allowing for interrogation of protein knockdown effects on in vivo neoangiogenesis in a Matrigel plug assay. In addition to interrogating the effect of gene knockdown in endothelial cells, we utilized lentiviral shRNA to knockdown specificity protein 1 (SP1), a transcription factor involved in the expression of VEGF, in U-87 MG tumor cells to demonstrate the ability to analyze angiogenesis in vitro in a tumor-driven transwell cord formation system and in tumor angiogenesis in vivo. A significant reduction in tumor-driven cord formation, VEGF secretion, and in vivo tumor angiogenesis was observed upon SP1 knockdown. Therefore, evaluation of target gene knockdown effects in the in vitro co-culture cord formation assay in the ADSC/ECFC co-culture, ECFCs alone, and in tumor cells translated directly to in vivo results, indicating the in vitro method as a robust, cost-effective and efficient in vitro surrogate assay to investigate target gene involvement in endothelial or tumor cell function in angiogenesis.
Conflict of interest statement
Figures

References
-
- Kebers F, Lewalle JM, Desreux J, Munaut C, Devy L, et al. (1998) Induction of endothelial cell apoptosis by solid tumor cells. Exp Cell Res 240: 197–205. - PubMed
-
- Bishop ET, Bell GT, Bloor S, Broom IJ, Hendry NF, et al. (1999) An in vitro model of angiogenesis: basic features. Angiogenesis 3: 335–344. - PubMed
-
- Jih YJ, Lien WH, Tsai WC, Yang GW, Li C, et al. (2001) Distinct regulation of genes by bFGF and VEGF-A in endothelial cells. Angiogenesis 4: 313–321. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
