

The interplay of seven subthreshold conductances controls the resting membrane potential and the oscillatory behavior of thalamocortical neurons
- PMID: 24760784
- PMCID: PMC4064413
- DOI: 10.1152/jn.00647.2013
The interplay of seven subthreshold conductances controls the resting membrane potential and the oscillatory behavior of thalamocortical neurons
Abstract
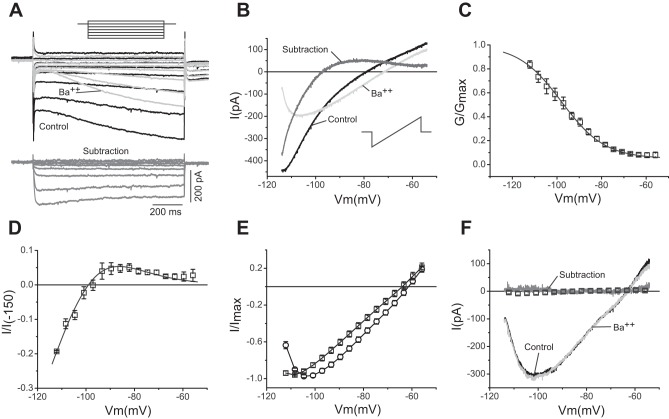
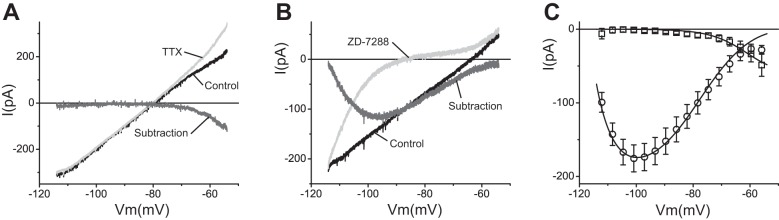
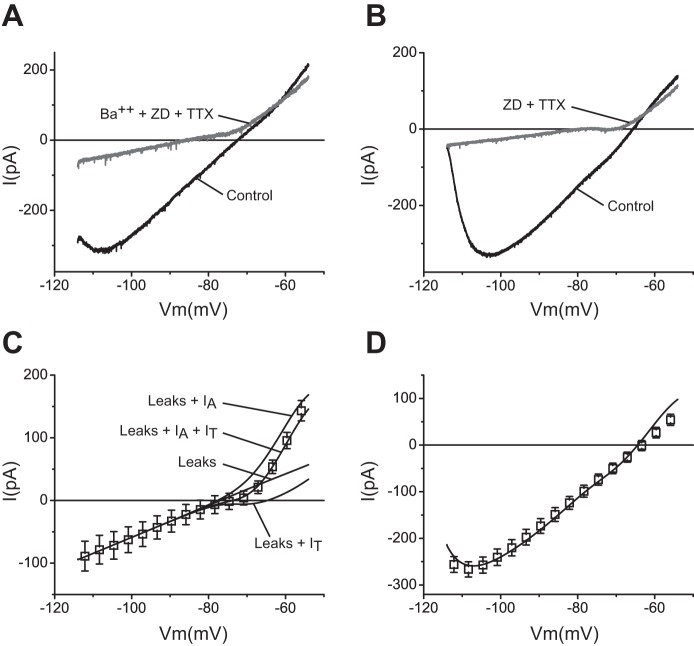
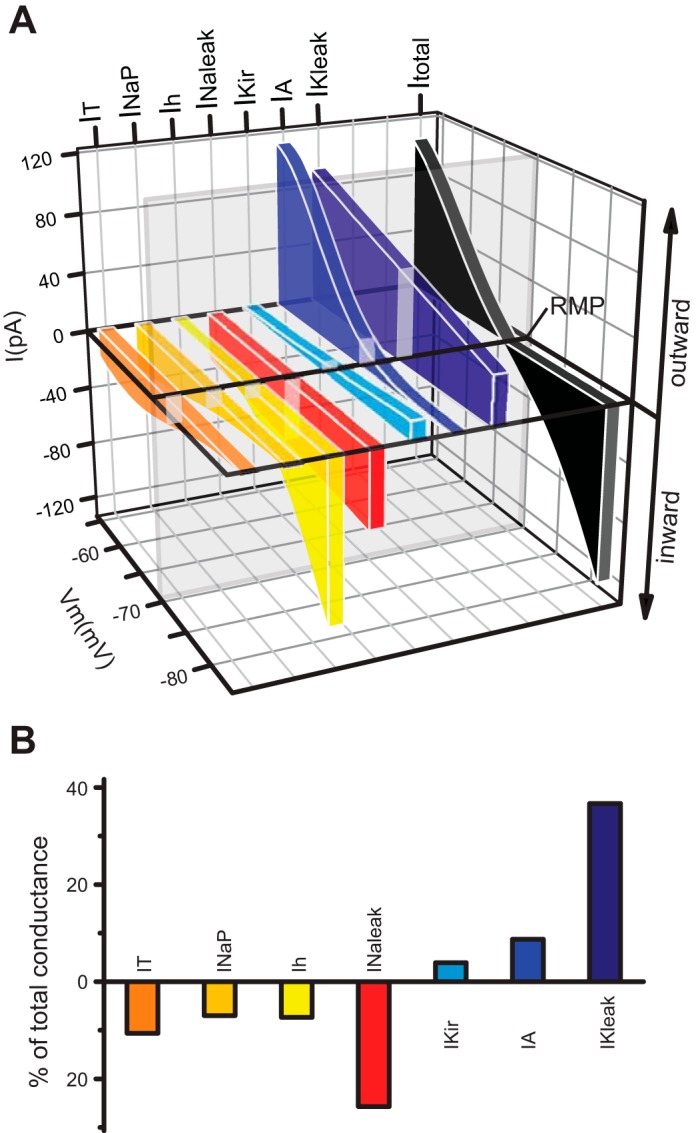
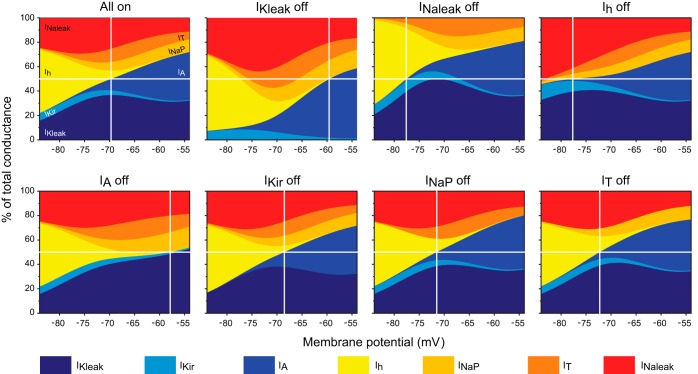
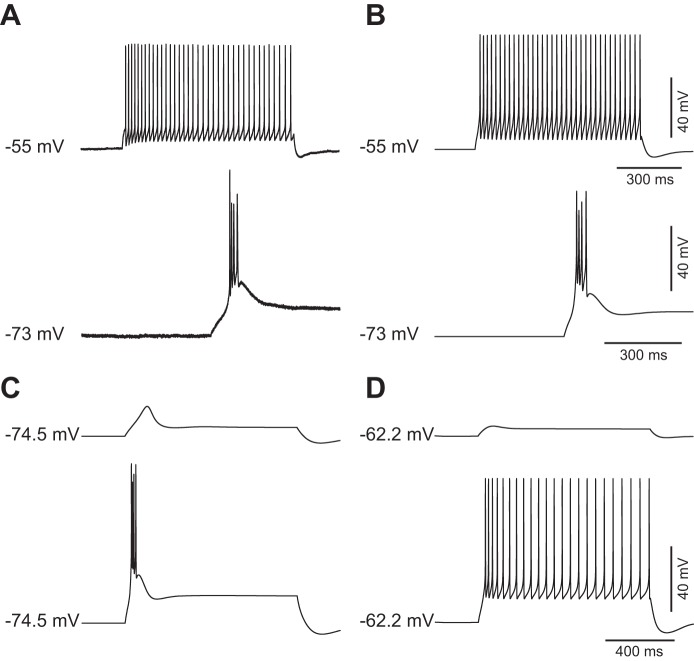
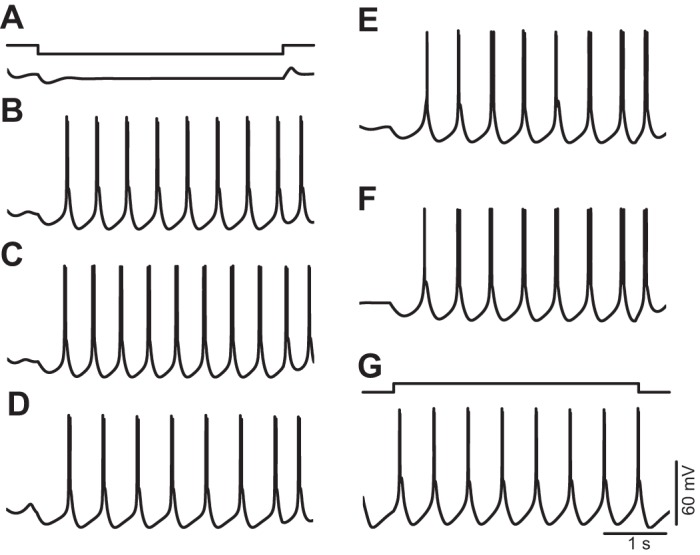
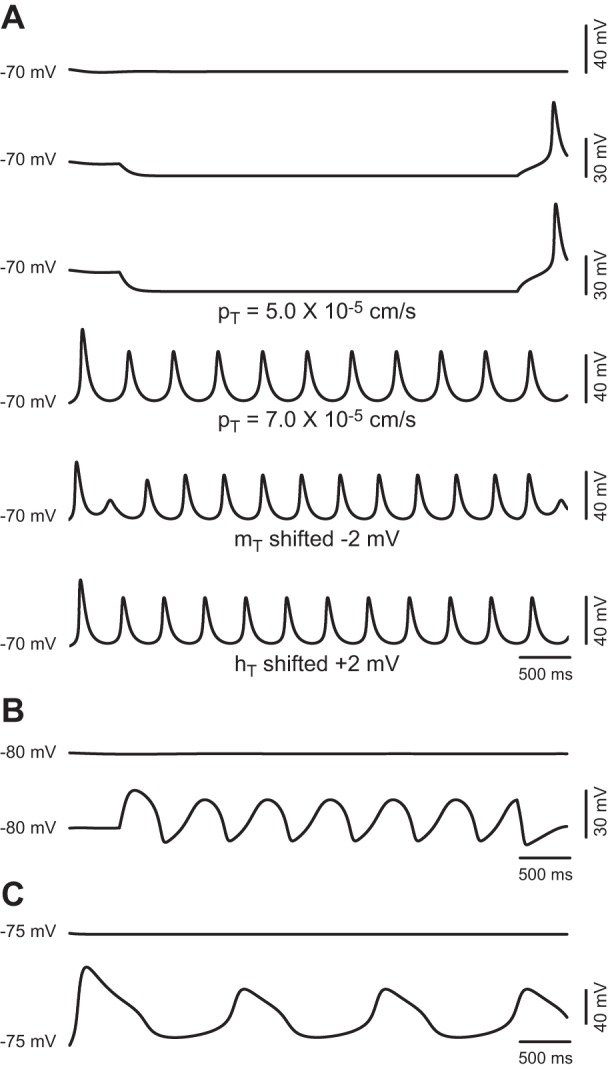
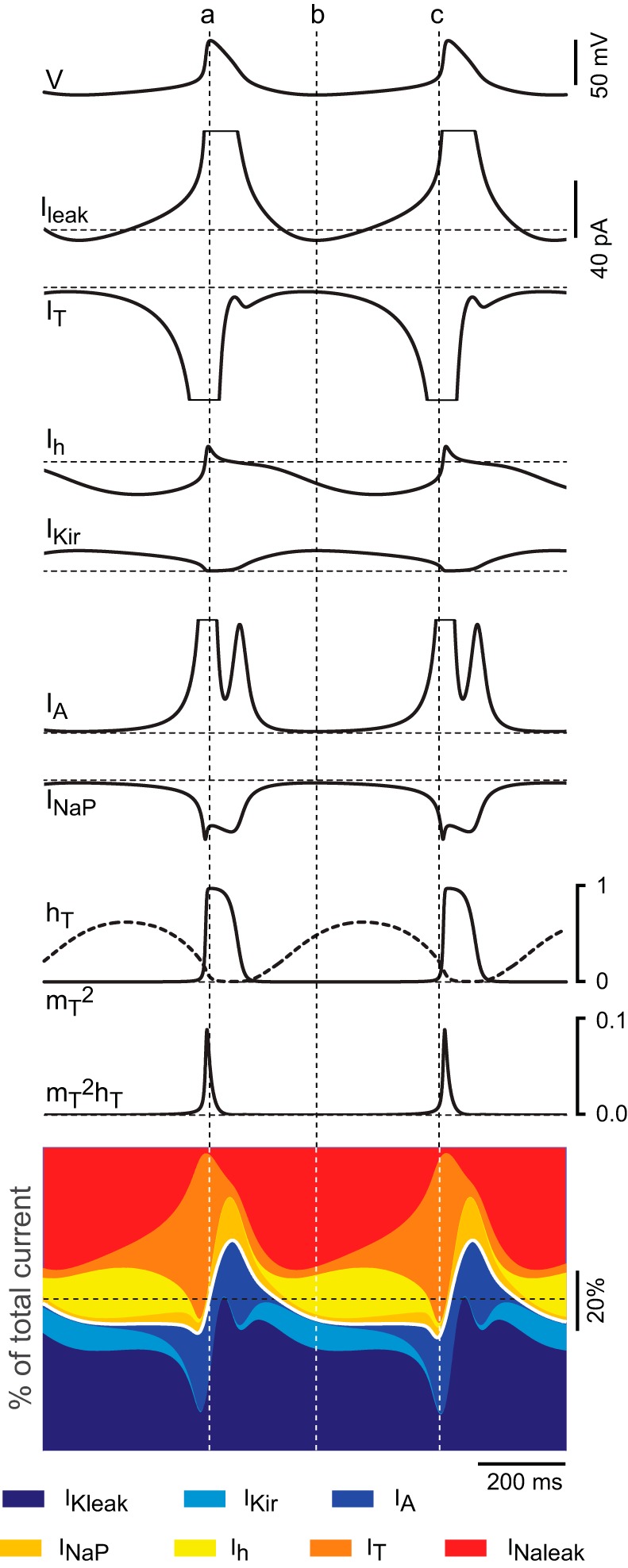
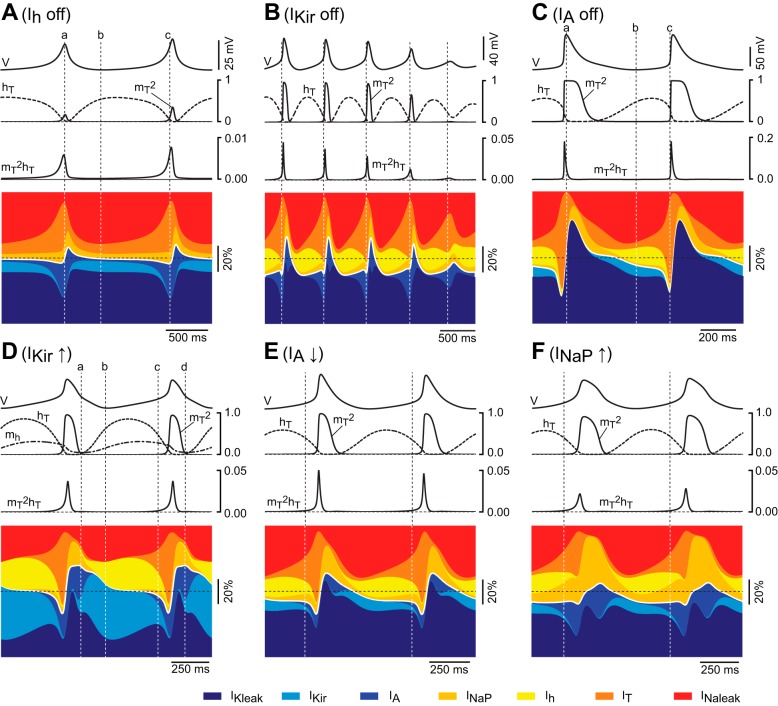
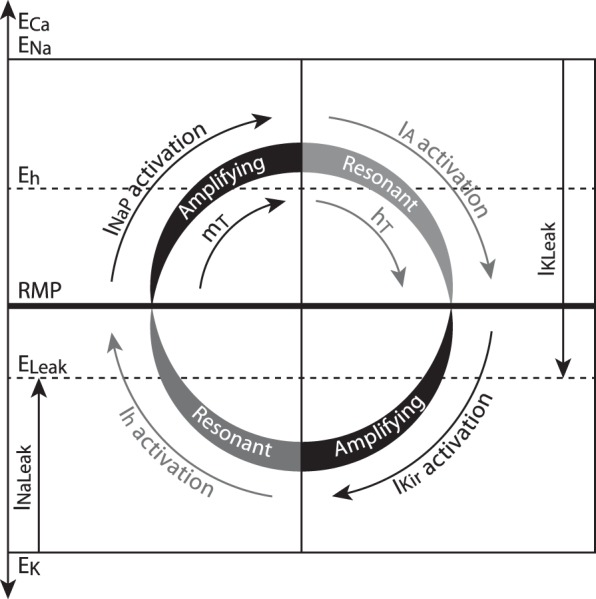
The signaling properties of thalamocortical (TC) neurons depend on the diversity of ion conductance mechanisms that underlie their rich membrane behavior at subthreshold potentials. Using patch-clamp recordings of TC neurons in brain slices from mice and a realistic conductance-based computational model, we characterized seven subthreshold ion currents of TC neurons and quantified their individual contributions to the total steady-state conductance at levels below tonic firing threshold. We then used the TC neuron model to show that the resting membrane potential results from the interplay of several inward and outward currents over a background provided by the potassium and sodium leak currents. The steady-state conductances of depolarizing Ih (hyperpolarization-activated cationic current), IT (low-threshold calcium current), and INaP (persistent sodium current) move the membrane potential away from the reversal potential of the leak conductances. This depolarization is counteracted in turn by the hyperpolarizing steady-state current of IA (fast transient A-type potassium current) and IKir (inwardly rectifying potassium current). Using the computational model, we have shown that single parameter variations compatible with physiological or pathological modulation promote burst firing periodicity. The balance between three amplifying variables (activation of IT, activation of INaP, and activation of IKir) and three recovering variables (inactivation of IT, activation of IA, and activation of Ih) determines the propensity, or lack thereof, of repetitive burst firing of TC neurons. We also have determined the specific roles that each of these variables have during the intrinsic oscillation.
Keywords: repetitive burst firing; resting membrane potential; subthreshold conductances; thalamocortical neuron.
Copyright © 2014 the American Physiological Society.
Figures
References
-
- Amarillo Y. Control of Neuronal Excitability by Subthreshold Operating Potassium Channels (PhD thesis). New York: New York University, 2007
-
- Amarillo Y, Nadal MS. On the repetitive burst firing of thalamic relay neurons. Program No. 46.08. 2011 Neuroscience Meeting Planner. Washington, DC: Society for Neuroscience, 2011
-
- Amarillo Y, Nadal MS, Rudy B. The inward rectifying potassium channel Kir2.2 regulates the excitability of thalamic relay neurons. Program No. 174.3 2005 Neuroscience Meeting Planner Washington, DC: Society for Neuroscience, 2005
-
- Biel M, Wahl-Schott C, Michalakis S, Zong X. Hyperpolarization-activated cation channels: from genes to function. Physiol Rev 89: 847–885, 2009 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources