

Nerve injury-induced neuropathic pain causes disinhibition of the anterior cingulate cortex
- PMID: 24760836
- PMCID: PMC6608297
- DOI: 10.1523/JNEUROSCI.3667-13.2014
Nerve injury-induced neuropathic pain causes disinhibition of the anterior cingulate cortex
Abstract
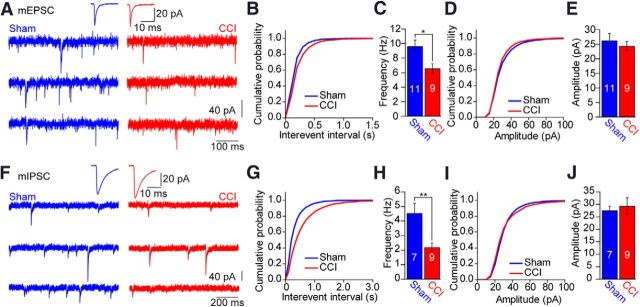
Neuropathic pain caused by peripheral nerve injury is a debilitating neurological condition of high clinical relevance. On the cellular level, the elevated pain sensitivity is induced by plasticity of neuronal function along the pain pathway. Changes in cortical areas involved in pain processing contribute to the development of neuropathic pain. Yet, it remains elusive which plasticity mechanisms occur in cortical circuits. We investigated the properties of neural networks in the anterior cingulate cortex (ACC), a brain region mediating affective responses to noxious stimuli. We performed multiple whole-cell recordings from neurons in layer 5 (L5) of the ACC of adult mice after chronic constriction injury of the sciatic nerve of the left hindpaw and observed a striking loss of connections between excitatory and inhibitory neurons in both directions. In contrast, no significant changes in synaptic efficacy in the remaining connected pairs were found. These changes were reflected on the network level by a decrease in the mEPSC and mIPSC frequency. Additionally, nerve injury resulted in a potentiation of the intrinsic excitability of pyramidal neurons, whereas the cellular properties of interneurons were unchanged. Our set of experimental parameters allowed constructing a neuronal network model of L5 in the ACC, revealing that the modification of inhibitory connectivity had the most profound effect on increased network activity. Thus, our combined experimental and modeling approach suggests that cortical disinhibition is a fundamental pathological modification associated with peripheral nerve damage. These changes at the cortical network level might therefore contribute to the neuropathic pain condition.
Keywords: anterior cingulate cortex; chronic pain; disinhibition; neuronal network; structural plasticity.
Figures







References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical