

Intrinsic TGF-β signaling promotes age-dependent CD8+ T cell polyfunctionality attrition
- PMID: 24762437
- PMCID: PMC4038564
- DOI: 10.1172/JCI70522
Intrinsic TGF-β signaling promotes age-dependent CD8+ T cell polyfunctionality attrition
Abstract
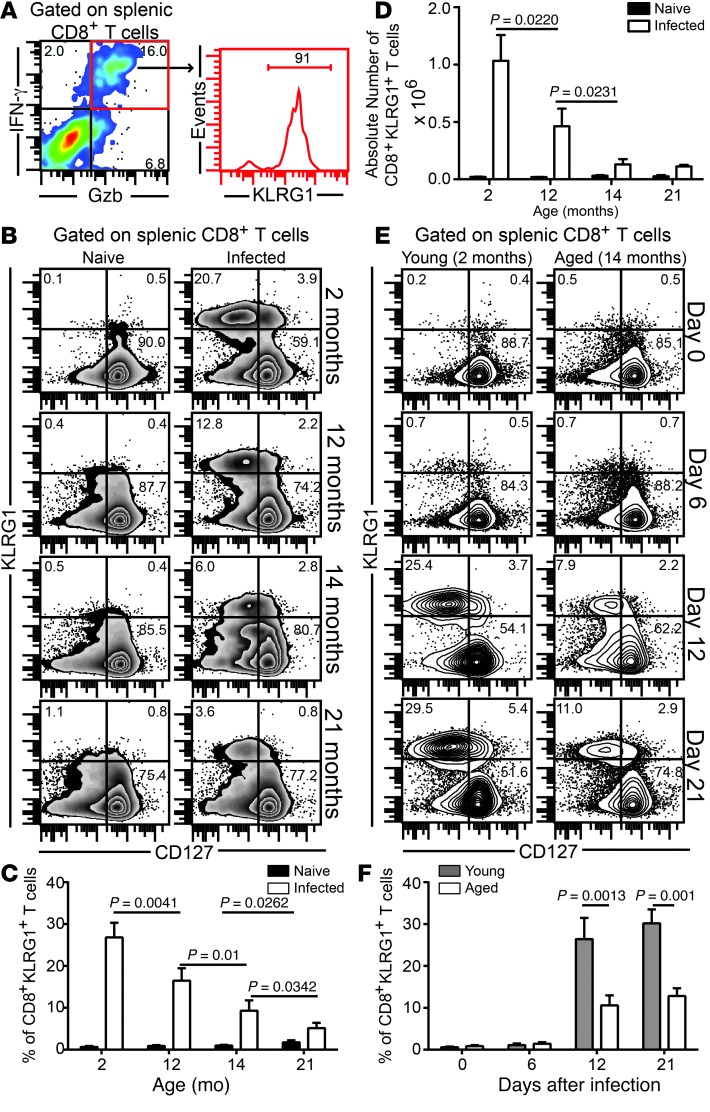
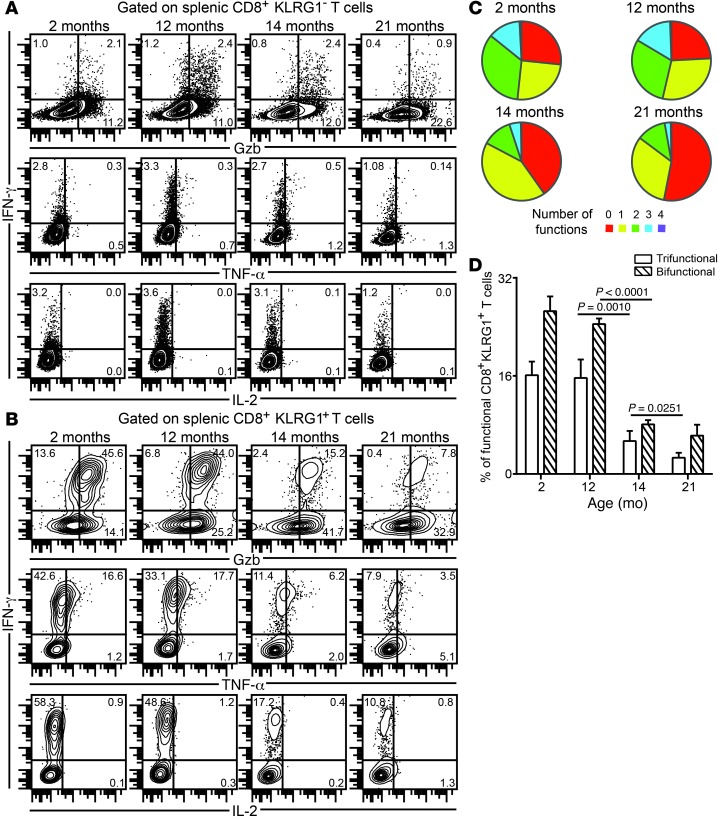
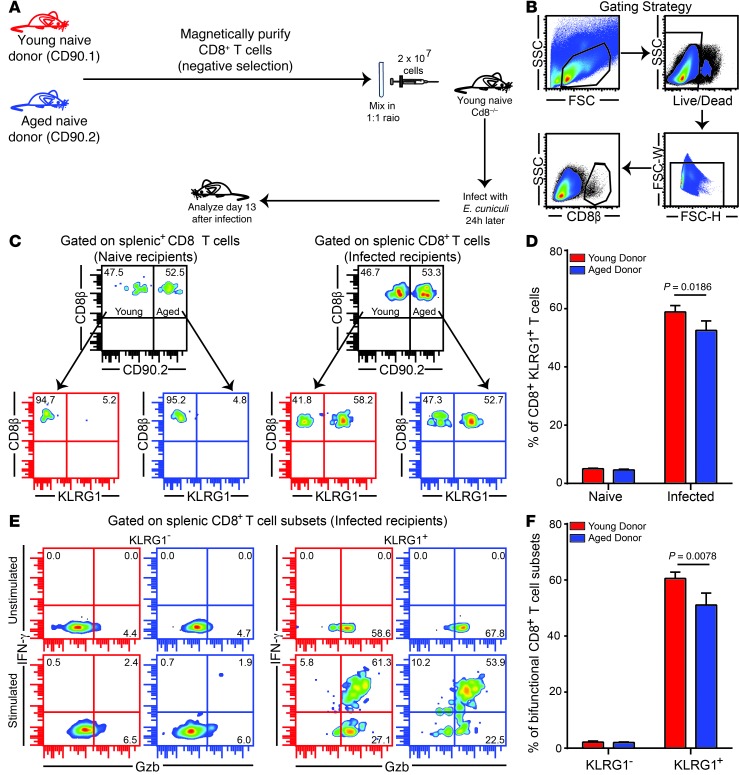
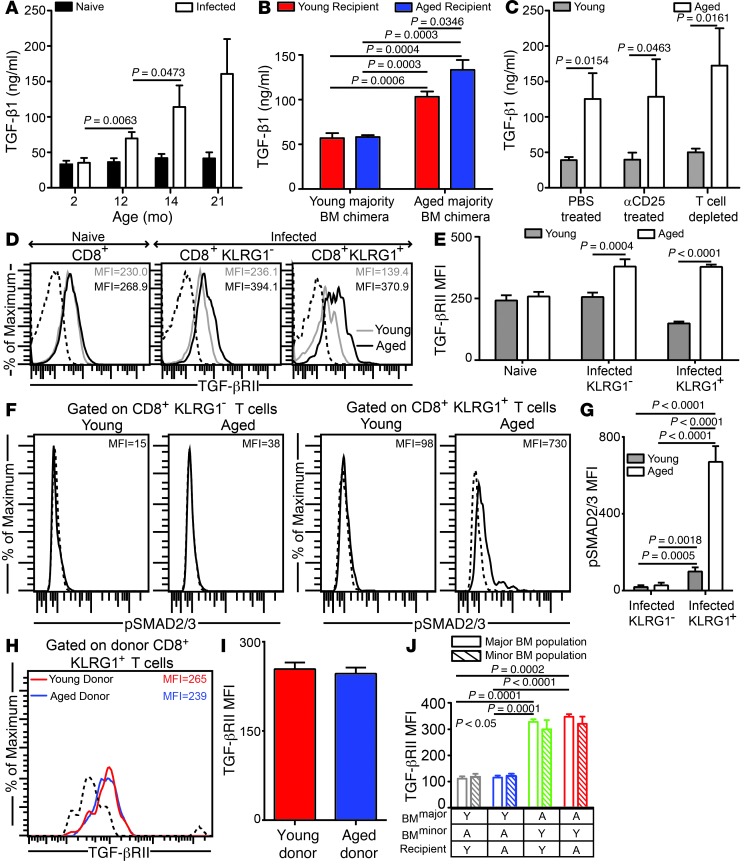
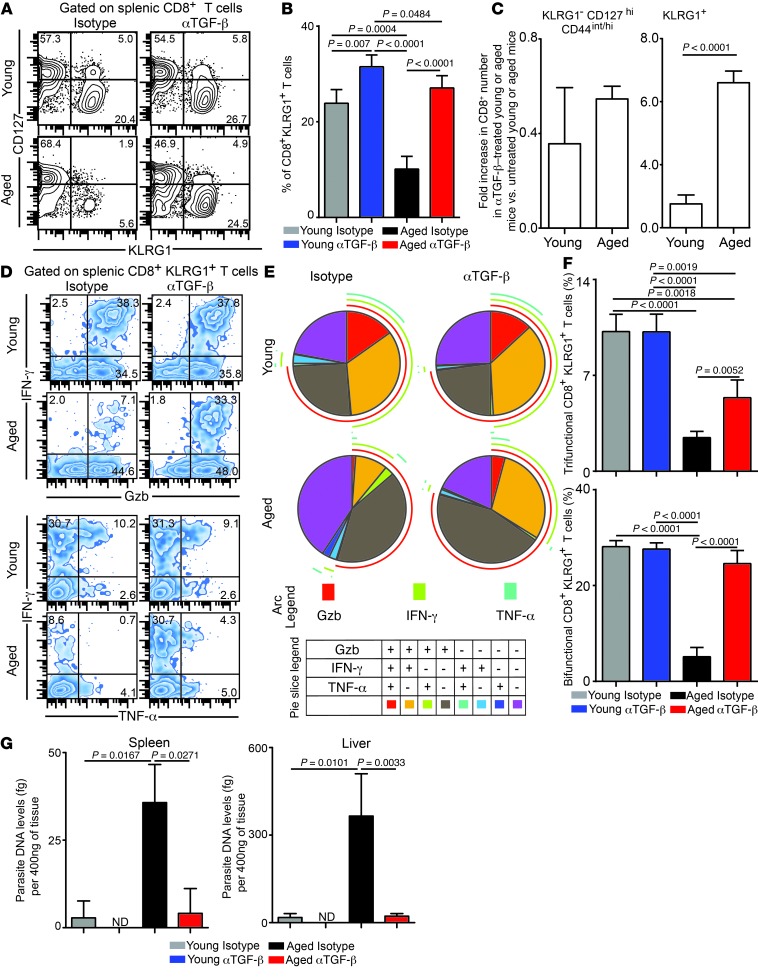
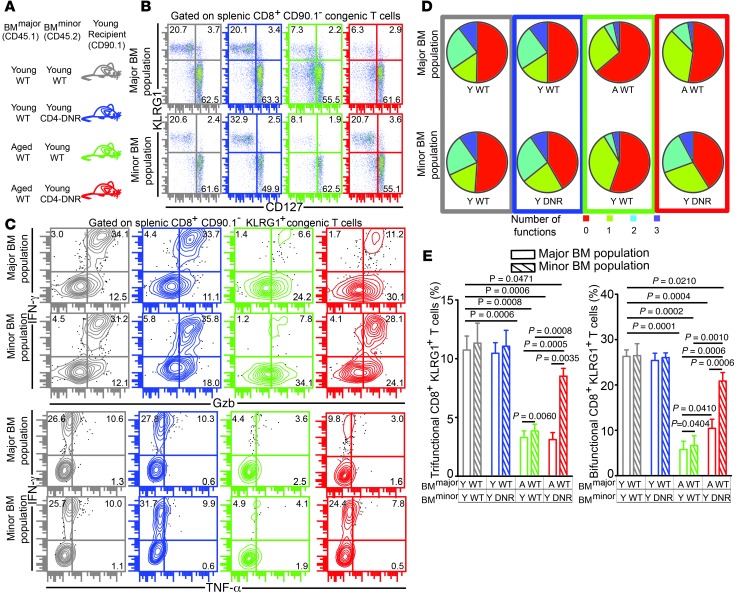
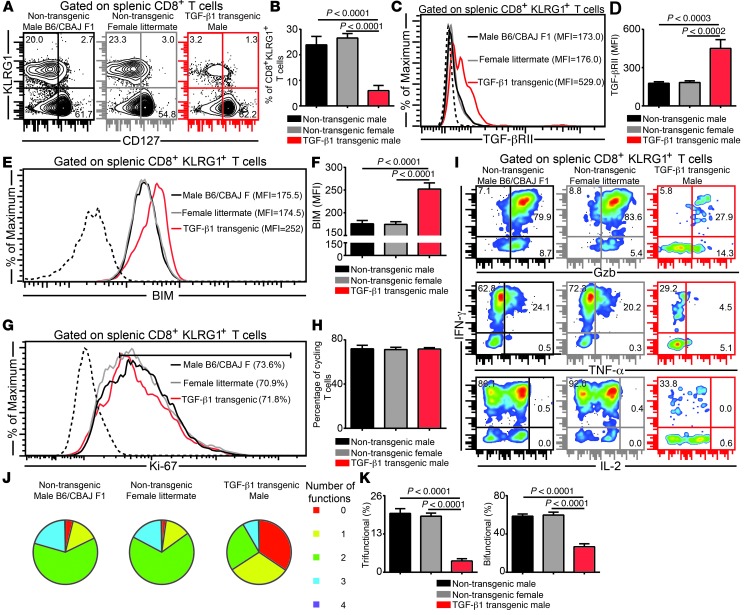
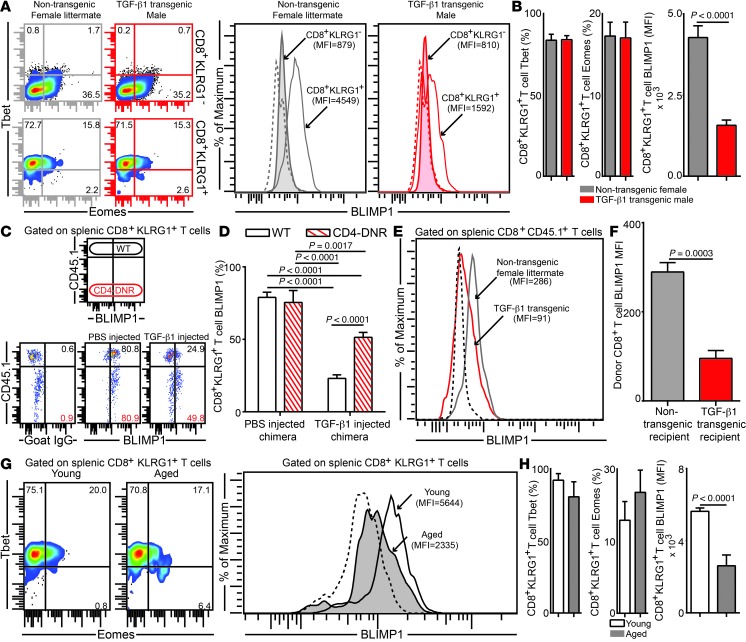
Advanced age is associated with immune system deficits that result in an increased susceptibility to infectious diseases; however, specific mediators of age-dependent immune dysfunction have not been fully elucidated. Here we demonstrated that aged mice exhibit poor effector CD8+ T cell polyfunctionality, primarily due to CD8+ T cell-extrinsic deficits, and that reduced CD8+ T cell polyfunctionality correlates with increased susceptibility to pathogenic diseases. In aged animals challenged with the parasite Encephalitozoon cuniculi, effector CD8+ T cell survival and polyfunctionality were suppressed by highly elevated TGF-β1. Furthermore, TGF-β depletion reduced effector CD8+ T cell apoptosis in both young and aged mice and enhanced effector CD8+ T cell polyfunctionality in aged mice. Surprisingly, intrinsic blockade of TGF-β signaling in CD8+ T cells was sufficient to rescue polyfunctionality in aged animals. Together, these data demonstrate that low levels of TGF-β1 promote apoptosis of CD8+ effector T cells and high TGF-β1 levels associated with age result in both CD8+ T cell apoptosis and an altered transcriptional profile, which correlates with loss of polyfunctionality. Furthermore, elevated TGF-β levels are observed in the elderly human population and in aged Drosophila, suggesting that TGF-β represents an evolutionarily conserved negative regulator of the immune response in aging organisms.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
