

Folliculin regulates ampk-dependent autophagy and metabolic stress survival
- PMID: 24763318
- PMCID: PMC3998892
- DOI: 10.1371/journal.pgen.1004273
Folliculin regulates ampk-dependent autophagy and metabolic stress survival
Abstract
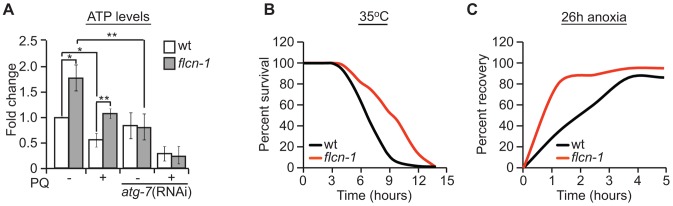
Dysregulation of AMPK signaling has been implicated in many human diseases, which emphasizes the importance of characterizing AMPK regulators. The tumor suppressor FLCN, responsible for the Birt-Hogg Dubé renal neoplasia syndrome (BHD), is an AMPK-binding partner but the genetic and functional links between FLCN and AMPK have not been established. Strikingly, the majority of naturally occurring FLCN mutations predisposing to BHD are predicted to produce truncated proteins unable to bind AMPK, pointing to the critical role of this interaction in the tumor suppression mechanism. Here, we demonstrate that FLCN is an evolutionarily conserved negative regulator of AMPK. Using Caenorhabditis elegans and mammalian cells, we show that loss of FLCN results in constitutive activation of AMPK which induces autophagy, inhibits apoptosis, improves cellular bioenergetics, and confers resistance to energy-depleting stresses including oxidative stress, heat, anoxia, and serum deprivation. We further show that AMPK activation conferred by FLCN loss is independent of the cellular energy state suggesting that FLCN controls the AMPK energy sensing ability. Together, our data suggest that FLCN is an evolutionarily conserved regulator of AMPK signaling that may act as a tumor suppressor by negatively regulating AMPK function.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures




References
-
- Hornstein OP, Knickenberg M (1975) Perifollicular fibromatosis cutis with polyps of the colon–a cutaneo-intestinal syndrome sui generis. Arch Dermatol Res 253: 161–175. - PubMed
-
- Birt AR, Hogg GR, Dube WJ (1977) Hereditary multiple fibrofolliculomas with trichodiscomas and acrochordons. Arch Dermatol 113: 1674–1677. - PubMed
-
- Toro JR, Glenn G, Duray P, Darling T, Weirich G, et al. (1999) Birt-Hogg-Dube syndrome: a novel marker of kidney neoplasia. Arch Dermatol 135: 1195–1202. - PubMed
-
- Pavlovich CP, Walther MM, Eyler RA, Hewitt SM, Zbar B, et al. (2002) Renal tumors in the Birt-Hogg-Dube syndrome. Am J Surg Pathol 26: 1542–1552. - PubMed
-
- Zbar B, Alvord WG, Glenn G, Turner M, Pavlovich CP, et al. (2002) Risk of renal and colonic neoplasms and spontaneous pneumothorax in the Birt-Hogg-Dube syndrome. Cancer Epidemiol Biomarkers Prev 11: 393–400. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
