

Mycobacterium indicus pranii and Mycobacterium bovis BCG lead to differential macrophage activation in Toll-like receptor-dependent manner
- PMID: 24766519
- PMCID: PMC4172141
- DOI: 10.1111/imm.12306
Mycobacterium indicus pranii and Mycobacterium bovis BCG lead to differential macrophage activation in Toll-like receptor-dependent manner
Abstract
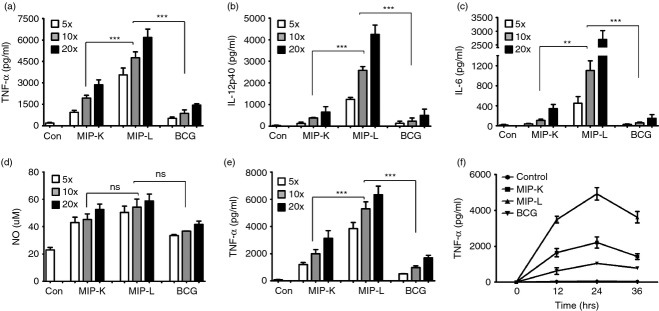
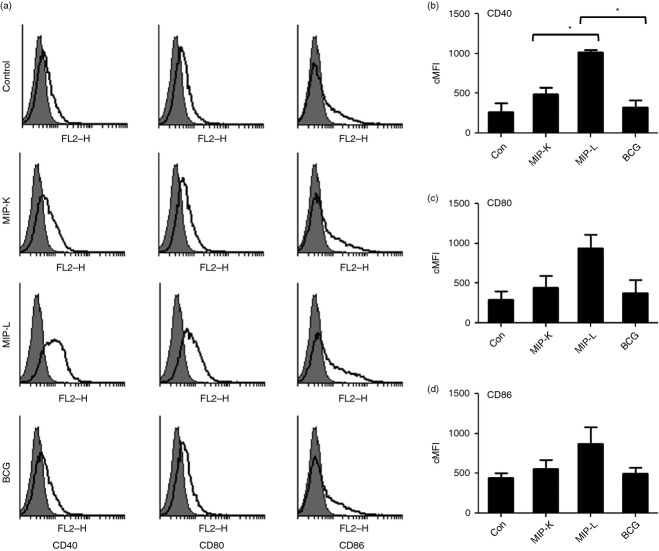
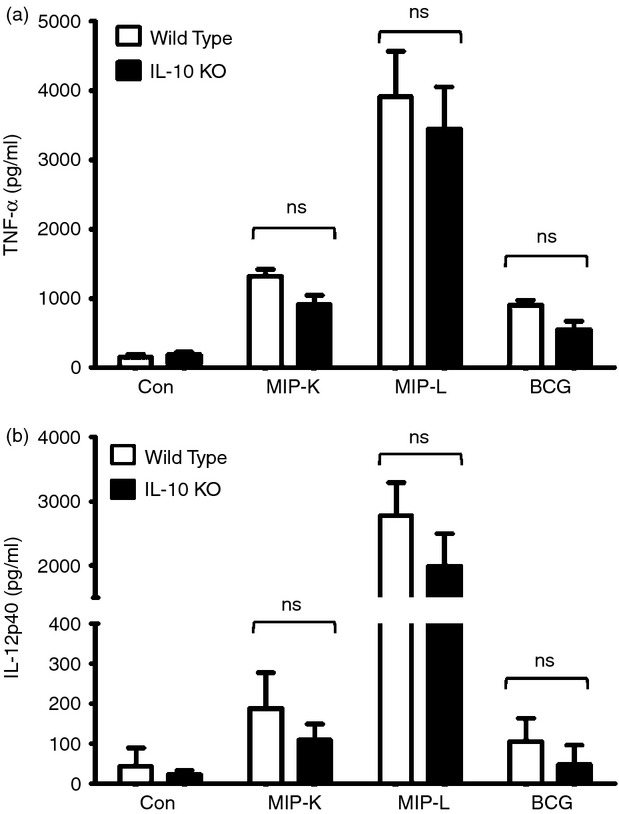
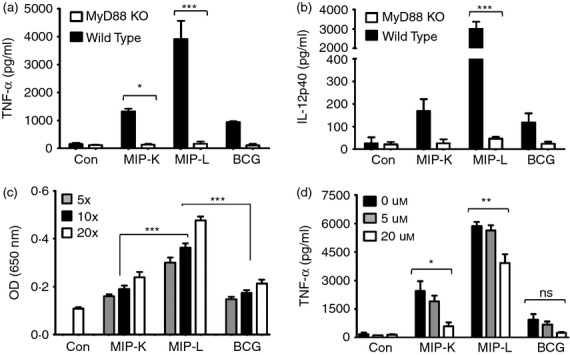
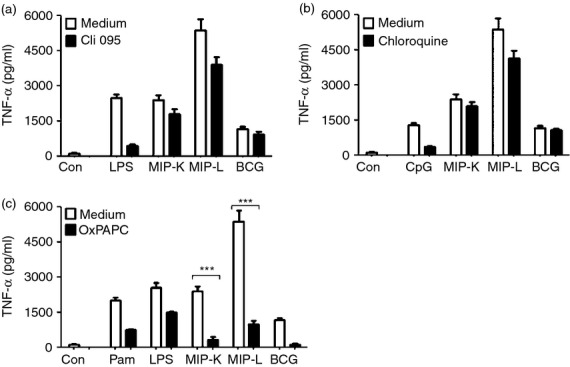
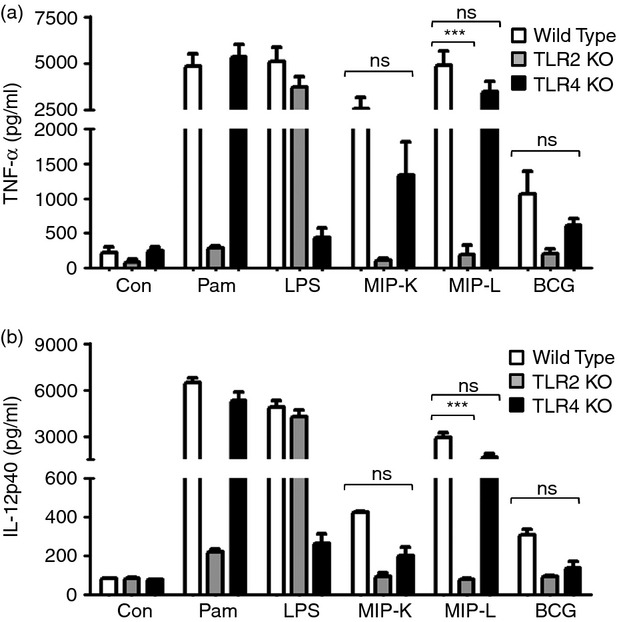
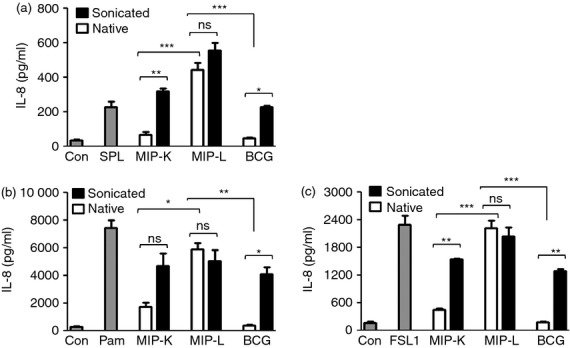
Mycobacterium indicus pranii (MIP) is an atypical mycobacterial species possessing strong immunomodulatory properties. It is a potent vaccine candidate against tuberculosis, promotes Th1 immune response and protects mice from tumours. In previous studies, we demonstrated higher protective efficacy of MIP against experimental tuberculosis as compared with bacillus Calmette-Guérin (BCG). Since macrophages play an important role in the pathology of mycobacterial diseases and cancer, in the present study, we evaluated the MIP in live and killed form for macrophage activation potential, compared it with BCG and investigated the underlying mechanisms. High levels of tumour necrosis factor-α, interleukin-12p40 (IL-12p40), IL-6 and nitric oxide were produced by MIP-stimulated macrophages as compared with BCG-stimulated macrophages. Prominent up-regulation of co-stimulatory molecules CD40, CD80 and CD86 was also observed in response to MIP. Loss of response in MyD88-deficient macrophages showed that both MIP and BCG activate the macrophages in a MyD88-dependent manner. MyD88 signalling pathway culminates in nuclear factor-κB/activator protein-1 (NF-κB/AP-1) activation and higher activation of NF-κB/AP-1 was observed in response to MIP. With the help of pharmacological inhibitors and Toll-like receptor (TLR) -deficient macrophages, we observed the role of TLR2, TLR4 and intracellular TLRs in MIP-mediated macrophage activation. Stimulation of HEK293 cells expressing TLR2 in homodimeric or heterodimeric form showed that MIP has a distinctly higher level of TLR2 agonist activity compared with BCG. Further experiments suggested that TLR2 ligands are well exposed in MIP whereas they are obscured in BCG. Our findings establish the higher macrophage activation potential of MIP compared with BCG and delineate the underlying mechanism.
Keywords: Toll-like receptors; bacterial; cytokines.
© 2014 John Wiley & Sons Ltd.
Figures
References
-
- Saxena VK, Singh US, Singh AK. Bacteriological study of a rapidly growing strain of Mycobacterium. Lepr India. 1978;50:588–96. - PubMed
-
- Deo MG. Anti-leprosy vaccines – field trials and future prospects. Indian J Lepr. 1984;56:764–75. - PubMed
-
- Talwar GP, Zaheer SA, Mukherjee R, et al. Immunotherapeutic effects of a vaccine based on a saprophytic cultivable mycobacterium, Mycobacterium w in multibacillary leprosy patients. Vaccine. 1990;8:121–9. - PubMed
-
- Katoch K, Katoch VM, Natrajan M, et al. Treatment of bacilliferous BL/LL cases with combined chemotherapy and immunotherapy. Int J Lepr Other Mycobact Dis. 1995;63:202–12. - PubMed
-
- Sharma P, Misra RS, Kar HK, Mukherjee A, Poricha D, Kaur H, Mukherjee R, Rani R. Mycobacterium w vaccine, a useful adjuvant to multidrug therapy in multibacillary leprosy: a report on hospital based immunotherapeutic clinical trials with a follow-up of 1–7 years after treatment. Lepr Rev. 2000;71:179–92. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
