

Effect of advanced intercrossing on genome structure and on the power to detect linked quantitative trait loci in a multi-parent population: a simulation study in rice
- PMID: 24767139
- PMCID: PMC4101851
- DOI: 10.1186/1471-2156-15-50
Effect of advanced intercrossing on genome structure and on the power to detect linked quantitative trait loci in a multi-parent population: a simulation study in rice
Abstract
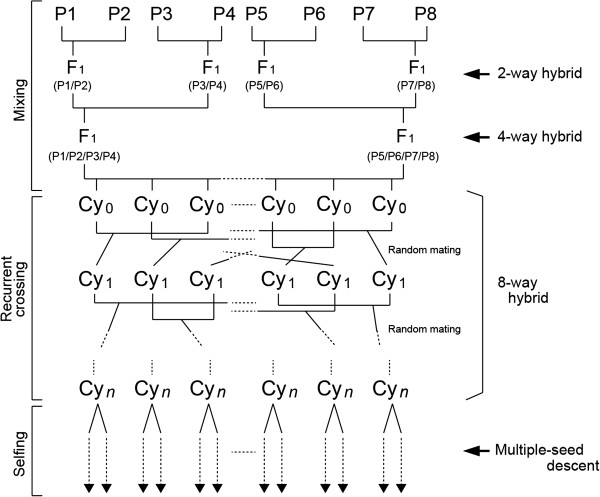
Background: In genetic analysis of agronomic traits, quantitative trait loci (QTLs) that control the same phenotype are often closely linked. Furthermore, many QTLs are localized in specific genomic regions (QTL clusters) that include naturally occurring allelic variations in different genes. Therefore, linkage among QTLs may complicate the detection of each individual QTL. This problem can be resolved by using populations that include many potential recombination sites. Recently, multi-parent populations have been developed and used for QTL analysis. However, their efficiency for detection of linked QTLs has not received attention. By using information on rice, we simulated the construction of a multi-parent population followed by cycles of recurrent crossing and inbreeding, and we investigated the resulting genome structure and its usefulness for detecting linked QTLs as a function of the number of cycles of recurrent crossing.
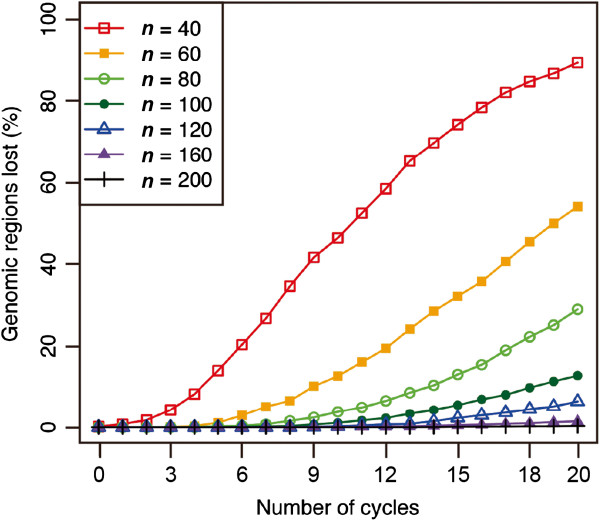
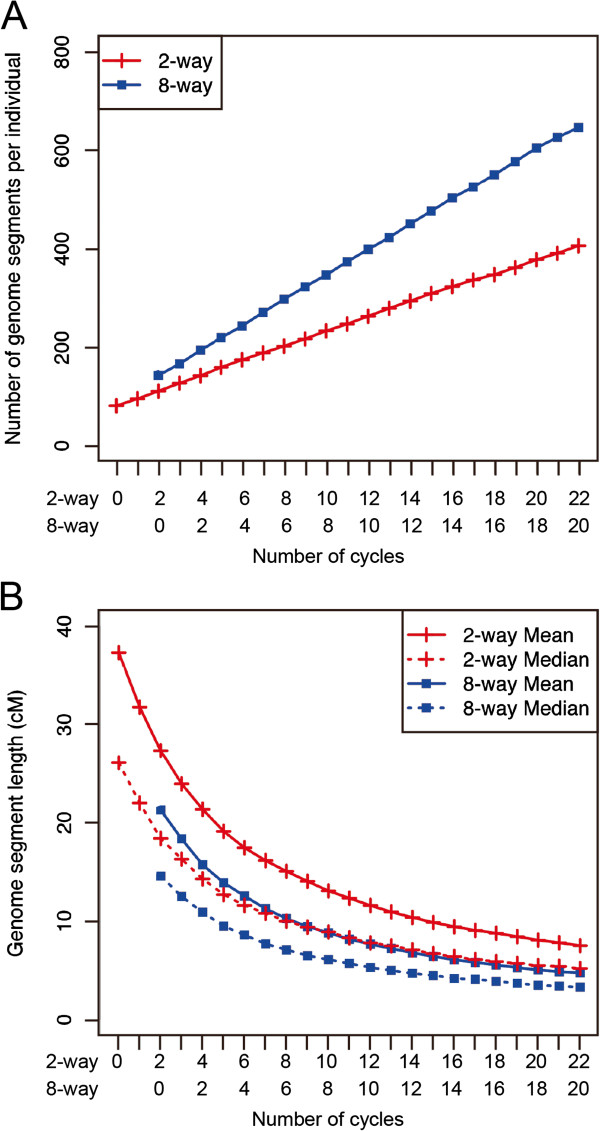
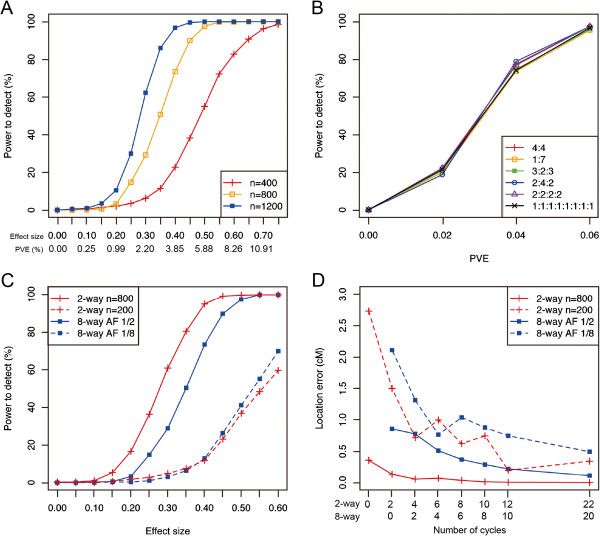
Results: The number of non-recombinant genome segments increased linearly with an increasing number of cycles. The mean and median lengths of the non-recombinant genome segments decreased dramatically during the first five to six cycles, then decreased more slowly during subsequent cycles. Without recurrent crossing, we found that there is a risk of missing QTLs that are linked in a repulsion phase, and a risk of identifying linked QTLs in a coupling phase as a single QTL, even when the population was derived from eight parental lines. In our simulation results, using fewer than two cycles of recurrent crossing produced results that differed little from the results with zero cycles, whereas using more than six cycles dramatically improved the power under most of the conditions that we simulated.
Conclusion: Our results indicated that even with a population derived from eight parental lines, fewer than two cycles of crossing does not improve the power to detect linked QTLs. However, using six cycles dramatically improved the power, suggesting that advanced intercrossing can help to resolve the problems that result from linkage among QTLs.
Figures
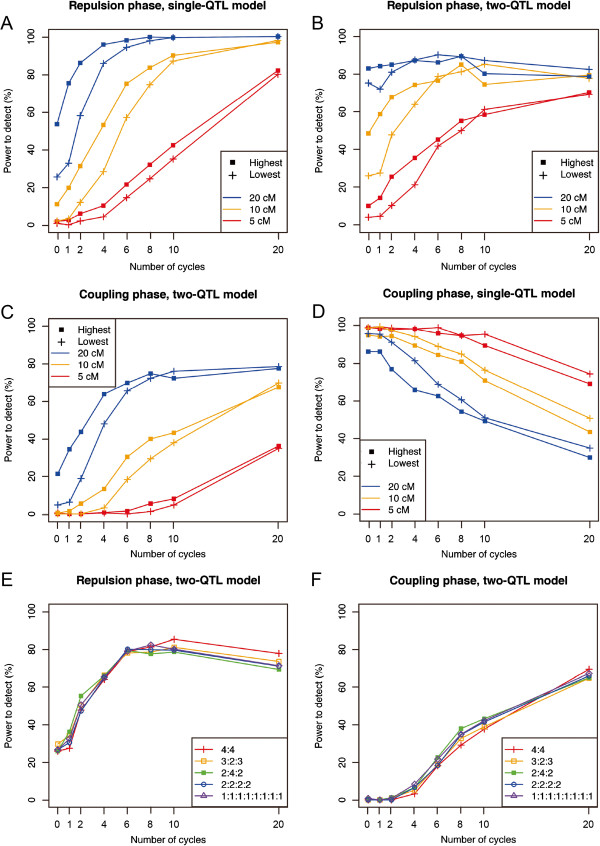
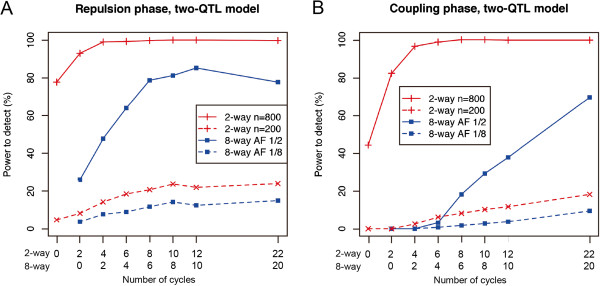
References
-
- Huang X, Wei X, Sang T, Zhao Q, Feng Q, Zhao Y, Li C, Zhu C, Lu T, Zhang Z, Li M, Fan D, Guo Y, Wang A, Wang L, Deng L, Li W, Lu Y, Weng Q, Liu K, Huang T, Zhou T, Jing Y, Li W, Lin Z, Buckler ES, Qian Q, Zhang QF, Li J, Han B. Genome-wide association studies of 14 agronomic traits in rice landraces. Nat Genet. 2010;42:961–967. doi: 10.1038/ng.695. - DOI - PubMed
-
- Huang X, Zhao Y, Wei X, Li C, Wang A, Zhao Q, Li W, Guo Y, Deng L, Zhu C, Fan D, Lu Y, Weng Q, Liu K, Zhou T, Jing Y, Si L, Dong G, Huang T, Lu T, Feng Q, Qian Q, Li J, Han B. Genome-wide association study of flowering time and grain yield traits in a worldwide collection of rice germplasm. Nat Genet. 2011;44:32–39. doi: 10.1038/ng.1018. - DOI - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
