

Stimulus-response bindings in priming
- PMID: 24768034
- PMCID: PMC4074350
- DOI: 10.1016/j.tics.2014.03.004
Stimulus-response bindings in priming
Abstract
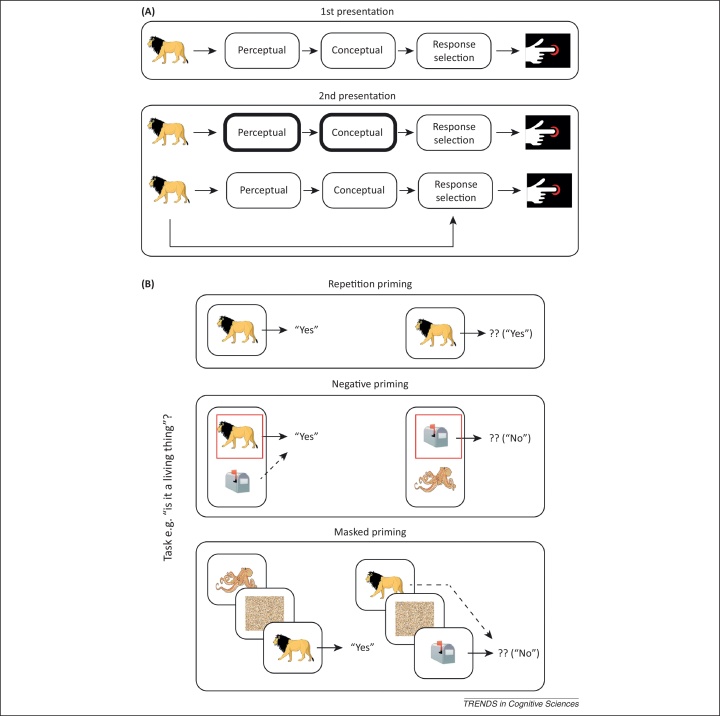
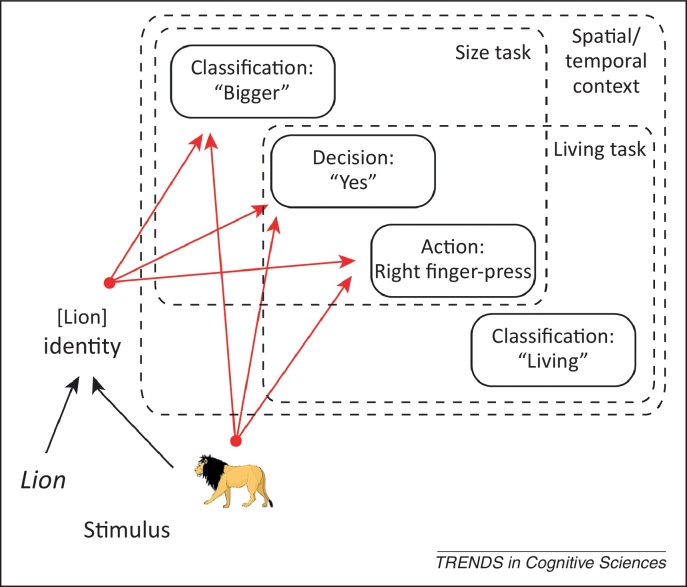
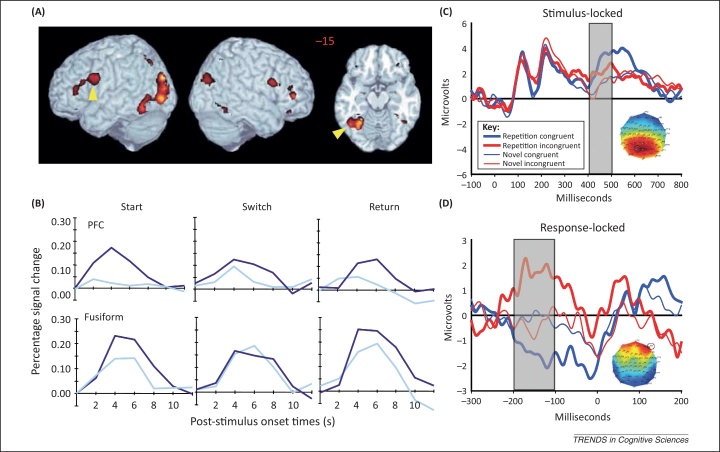
People can rapidly form arbitrary associations between stimuli and the responses they make in the presence of those stimuli. Such stimulus-response (S-R) bindings, when retrieved, affect the way that people respond to the same, or related, stimuli. Only recently, however, has the flexibility and ubiquity of these S-R bindings been appreciated, particularly in the context of priming paradigms. This is important for the many cognitive theories that appeal to evidence from priming. It is also important for the control of action generally. An S-R binding is more than a gradually learned association between a specific stimulus and a specific response; instead, it captures the full, context-dependent behavioral potential of a stimulus.
Keywords: S–R bindings; automaticity; masked priming; negative priming; repetition suppression; subliminal priming.
Copyright © 2014 The Authors. Published by Elsevier Ltd.. All rights reserved.
Figures
References
-
- Roediger H.L., III . Explaining dissociations between implicit and explicit measures of retention: a processing account. In: Roediger H.L. III, Craik F.I.M., editors. Varieties of Memory and Consciousness: Essays in Honor of Endel Tulving. Erlbaum; 1989. pp. 67–84.
-
- Logan G.D. Repetition priming and automaticity: common underlying mechanisms? Cogn. Psychol. 1990;22:1–35.
-
- Rothermund K. Retrieval of incidental stimulus–response associations as a source of negative priming. J. Exp. Psychol. Learn. 2005;31:482–495. - PubMed
-
- Damian M.F. Congruity effects evoked by subliminally presented primes: automaticity rather than semantic processing. J. Exp. Psychol. Hum. Percept. Perform. 2001;27:154–165. - PubMed
-
- Tipper S.P. Does negative priming reflect inhibitory mechanisms? A review and integration of conflicting views. Q. J. Exp. Psychol. (Colchester) 2001;54A:321–343. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
