

Candidate neural substrates for off-edge motion detection in Drosophila
- PMID: 24768048
- PMCID: PMC4031294
- DOI: 10.1016/j.cub.2014.03.051
Candidate neural substrates for off-edge motion detection in Drosophila
Abstract
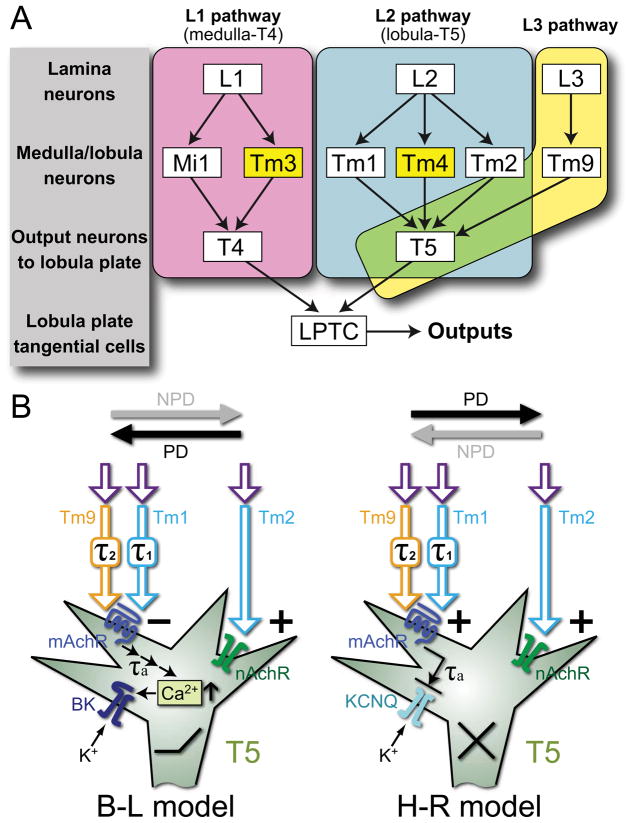
Background: In the fly's visual motion pathways, two cell types-T4 and T5-are the first known relay neurons to signal small-field direction-selective motion responses [1]. These cells then feed into large tangential cells that signal wide-field motion. Recent studies have identified two types of columnar neurons in the second neuropil, or medulla, that relay input to T4 from L1, the ON-channel neuron in the first neuropil, or lamina, thus providing a candidate substrate for the elementary motion detector (EMD) [2]. Interneurons relaying the OFF channel from L1's partner, L2, to T5 are so far not known, however.
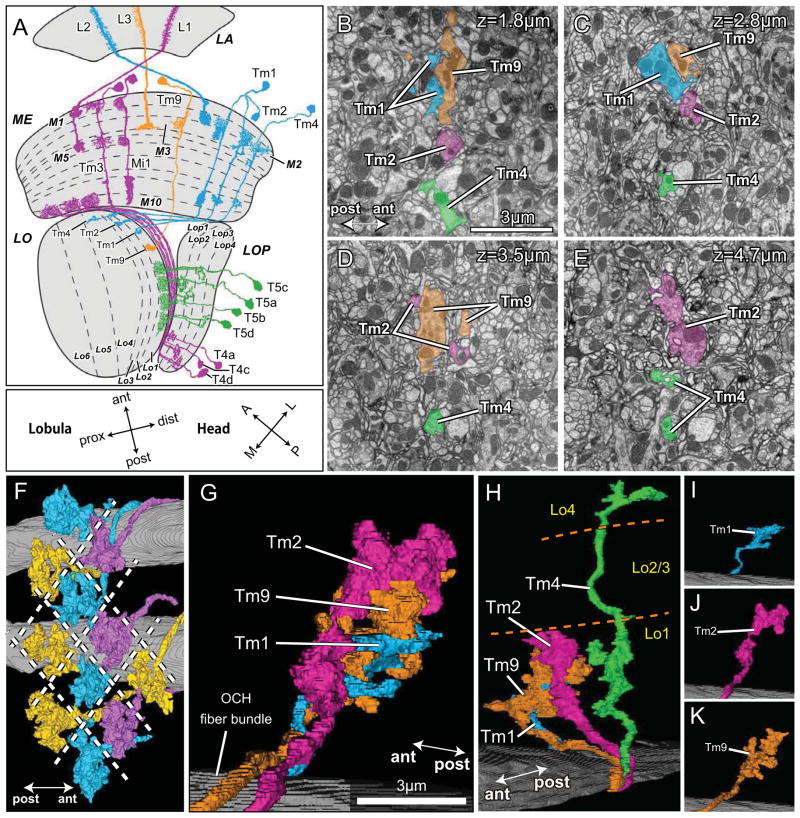
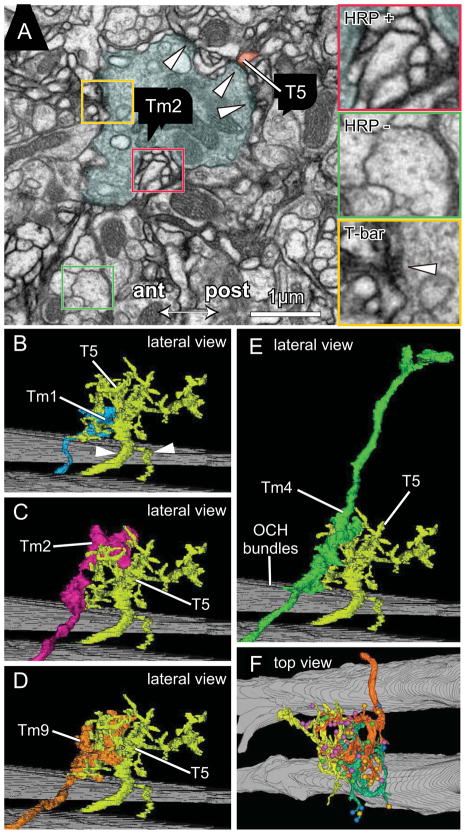
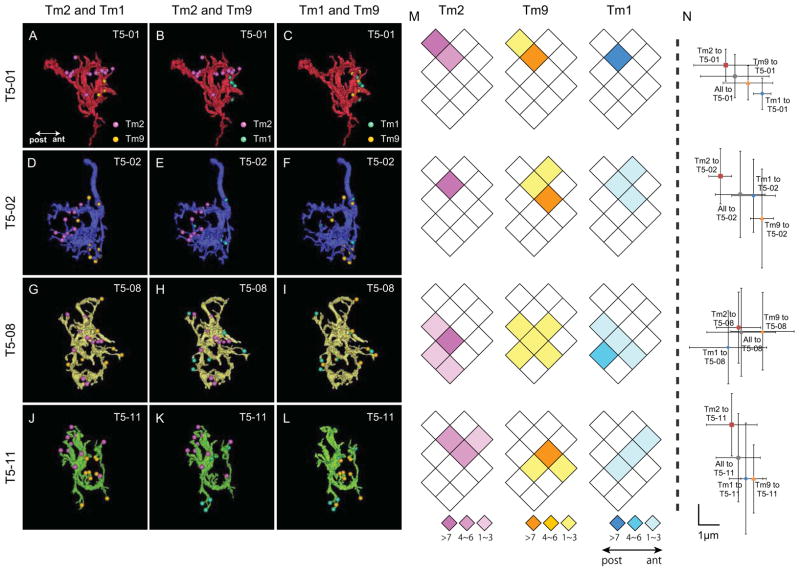
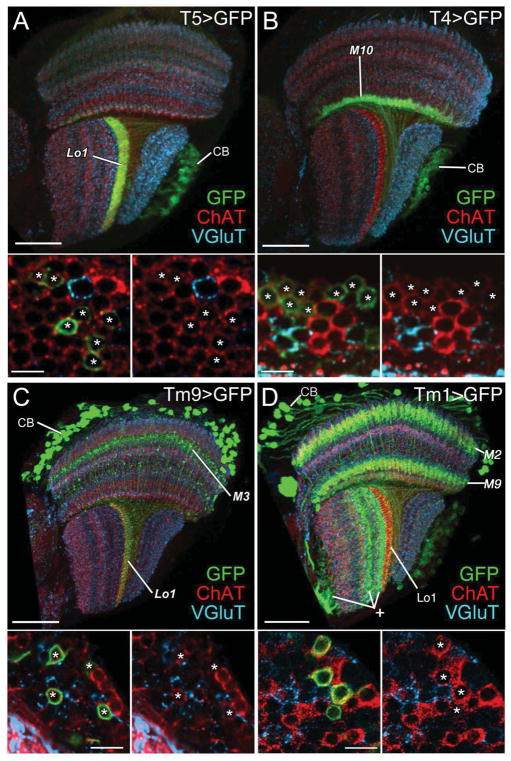
Results: Here we report that multiple types of transmedulla (Tm) neurons provide unexpectedly complex inputs to T5 at their terminals in the third neuropil, or lobula. From the L2 pathway, single-column input comes from Tm1 and Tm2 and multiple-column input from Tm4 cells. Additional input to T5 comes from Tm9, the medulla target of a third lamina interneuron, L3, providing a candidate substrate for L3's combinatorial action with L2 [3]. Most numerous, Tm2 and Tm9's input synapses are spatially segregated on T5's dendritic arbor, providing candidate anatomical substrates for the two arms of a T5 EMD circuit; Tm1 and Tm2 provide a second. Transcript profiling indicates that T5 expresses both nicotinic and muscarinic cholinoceptors, qualifying T5 to receive cholinergic inputs from Tm9 and Tm2, which both express choline acetyltransferase (ChAT).
Conclusions: We hypothesize that T5 computes small-field motion signals by integrating multiple cholinergic Tm inputs using nicotinic and muscarinic cholinoceptors.
Copyright © 2014 Elsevier Ltd. All rights reserved.
Figures
Comment in
-
Vision: EM-erging motion-detecting circuits.Curr Biol. 2014 May 19;24(10):R390-2. doi: 10.1016/j.cub.2014.03.067. Curr Biol. 2014. PMID: 24845666
Similar articles
-
How fly neurons compute the direction of visual motion.J Comp Physiol A Neuroethol Sens Neural Behav Physiol. 2020 Mar;206(2):109-124. doi: 10.1007/s00359-019-01375-9. Epub 2019 Nov 5. J Comp Physiol A Neuroethol Sens Neural Behav Physiol. 2020. PMID: 31691093 Free PMC article. Review.
-
A common evolutionary origin for the ON- and OFF-edge motion detection pathways of the Drosophila visual system.Front Neural Circuits. 2015 Jul 9;9:33. doi: 10.3389/fncir.2015.00033. eCollection 2015. Front Neural Circuits. 2015. PMID: 26217193 Free PMC article.
-
Cholinergic circuits integrate neighboring visual signals in a Drosophila motion detection pathway.Curr Biol. 2011 Dec 20;21(24):2077-84. doi: 10.1016/j.cub.2011.10.053. Epub 2011 Dec 1. Curr Biol. 2011. PMID: 22137471 Free PMC article.
-
Columnar cholinergic neurotransmission onto T5 cells of Drosophila.Curr Biol. 2025 Mar 24;35(6):1269-1284.e6. doi: 10.1016/j.cub.2025.02.004. Epub 2025 Feb 27. Curr Biol. 2025. PMID: 40020661
-
In search of the Holy Grail of fly motion vision.Eur J Neurosci. 2014 Nov;40(9):3285-93. doi: 10.1111/ejn.12731. Epub 2014 Sep 23. Eur J Neurosci. 2014. PMID: 25251169 Review.
Cited by
-
Processing properties of ON and OFF pathways for Drosophila motion detection.Nature. 2014 Aug 28;512(7515):427-30. doi: 10.1038/nature13427. Epub 2014 Jul 6. Nature. 2014. PMID: 25043016 Free PMC article.
-
Direct Measurement of Correlation Responses in Drosophila Elementary Motion Detectors Reveals Fast Timescale Tuning.Neuron. 2016 Oct 5;92(1):227-239. doi: 10.1016/j.neuron.2016.09.017. Neuron. 2016. PMID: 27710784 Free PMC article.
-
The Neuronal Basis of an Illusory Motion Percept Is Explained by Decorrelation of Parallel Motion Pathways.Curr Biol. 2018 Dec 3;28(23):3748-3762.e8. doi: 10.1016/j.cub.2018.10.007. Epub 2018 Nov 21. Curr Biol. 2018. PMID: 30471993 Free PMC article.
-
How fly neurons compute the direction of visual motion.J Comp Physiol A Neuroethol Sens Neural Behav Physiol. 2020 Mar;206(2):109-124. doi: 10.1007/s00359-019-01375-9. Epub 2019 Nov 5. J Comp Physiol A Neuroethol Sens Neural Behav Physiol. 2020. PMID: 31691093 Free PMC article. Review.
-
Common circuit design in fly and mammalian motion vision.Nat Neurosci. 2015 Aug;18(8):1067-76. doi: 10.1038/nn.4050. Epub 2015 Jun 29. Nat Neurosci. 2015. PMID: 26120965 Review.
References
-
- Maisak MS, Haag J, Ammer G, Serbe E, Meier M, Leonhardt A, Schilling T, Bahl A, Rubin GM, Nern A, et al. A directional tuning map of Drosophila elementary motion detectors. Nature. 2013;500:212–216. - PubMed
-
- Hassenstein B, Reichardt W. Systemtheoretische Analyse der Zeit-, Reihenfolgen- und Vorzeichenauswertung bei der Bewegungsperzeption des Rüsselkäfers Chlorophanus. Z Naturforsch. 1956;11:513–524.
-
- Reichardt W. Autocorrelation, a principle for the evaluation of sensory information by the central nervous system. In: Rosenblith WA, editor. In Sensory communication. New York/London: MIT Press; 1961. pp. 303–317.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
