

Review
doi: 10.1016/j.dnarep.2014.03.018.
Epub 2014 Apr 24.
Mechanism and regulation of incisions during DNA interstrand cross-link repair
Affiliations
- PMID: 24768452
- PMCID: PMC4076290
- DOI: 10.1016/j.dnarep.2014.03.018
Item in Clipboard
Review
Mechanism and regulation of incisions during DNA interstrand cross-link repair
DNA Repair (Amst).
2014 Jul.
Abstract
A critical step in DNA interstrand cross-link repair is the programmed collapse of replication forks that have stalled at an ICL. This event is regulated by the Fanconi anemia pathway, which suppresses bone marrow failure and cancer. In this perspective, we focus on the structure of forks that have stalled at ICLs, how these structures might be incised by endonucleases, and how incision is regulated by the Fanconi anemia pathway.
Keywords: DNA interstrand crosslink repair; EME1; ERCC1; FAN1; FANCD2; FANCI; Fanconi anemia; MUS81; SLX1; SLX4; SNM1A; XPF.
Copyright © 2014 Elsevier B.V. All rights reserved.
Figures
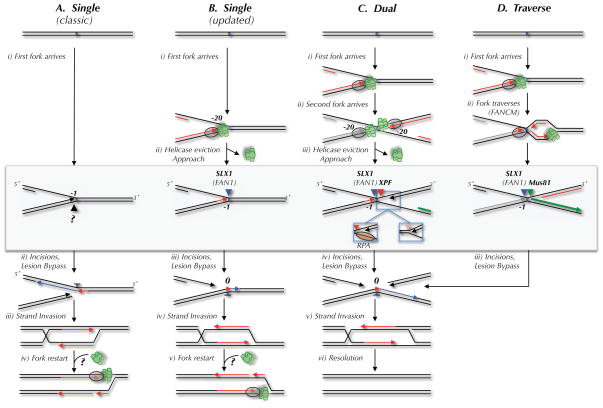
Four mechanisms of replication-dependent ICL repair are depicted. The DNA structures acted on by endonucleases in each model are highlighted by a gray box. Incisions are represented by black, blue, red, and green arrowheads. The proposed nuclease(s) that performs each incision is indicated above the arrowhead. (A) The classic ICL repair model, in which a single replication fork collides with the ICL and the leading strand template is incised (Niedernhofer et al., 2005). (B) The classic model, but taking into account the observation that leading strands initially stall 20 nucleotides from the ICL due to the MCM2-7 complex, and that incision occurs on the lagging strand template (Raschle et al., 2008). In models A and B, fork restart would require reloading of the MCM2-7 complex, for which there is no known mechanism. (C) The dual fork convergence model (Raschle et al., 2008). Left inset, 3′ incision substrate if RPA binds the lagging strand template after MCM2-7 removal. Right inset, 3′ incision substrate if parental strands re-anneal after MCM2-7 removal. (D) Traverse model (Huang et al., 2013). The only difference in the incision substrate in the dual fork and traverse models is the location of the 5′ end of the nascent strand on the right side of the ICL (green strand).
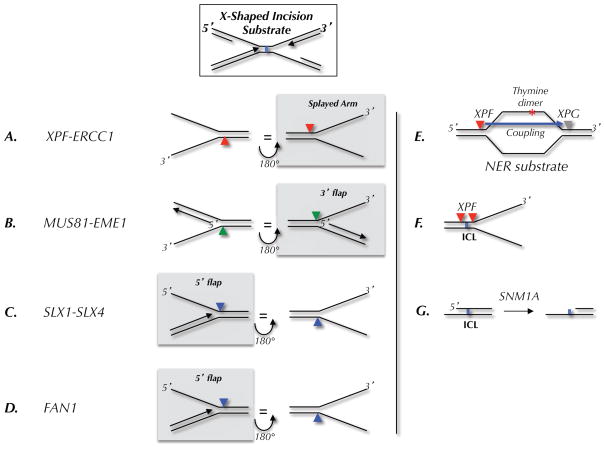
(A–D) Preferred substrates, as determined in biochemical assays, for the indicated endonucleases. The arrowheads indicate the location of incision. Each substrate is shown in two orientations related by a 180 degree rotation. The left column shows where the nuclease would cut on the 5′ side of a converged fork; the right column shows incisions on the 3′ side. In each column, the nucleases that are likely to cut the ICL-associated X-shaped structure shown at the top are highlighted by a grey box. (E) Classic nucleotide excision repair (NER) substrate and locations of XPF and XPG incisions. (F) Location of incisions when XPF is presented with an ICL-containing fork-shaped structure. (G) Action of SNM1A on ICL-containing DNA.
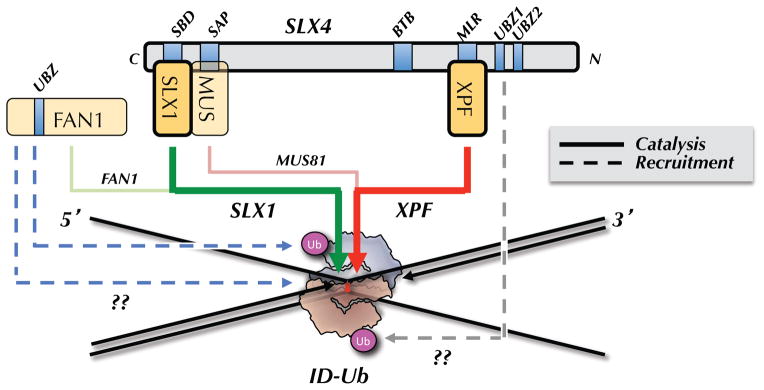
ID-Ub binds directly to an ICL in such a manner as to sequester the bottom parental strand while exposing the top parental strand. ID-Ub also recruits the SLX4-SLX1-MUS81-XPF-ERCC1 complex to the ICL, possibly via a direct interaction between the UBZ domains of SLX4 and the ubiquitin of the ID complex (dashed, grey arrow). XPF is the primary enzyme that incises 3′ to the ICL (thick red arrow), while SLX1 promotes the 5′ incision (green arrow). MUS81 performs a specialty role, cutting on the 3′ side in cases where the 5′ end of the leading strand on the same side abuts the ICL (pink arrow; shown in Figure 1D and 2B). FAN1 is recruited to ICLs via two independent mechanisms, one of which requires ID-Ub (dashed blue arrows). FAN1 cuts on the 5′ side of the ICL (light green arrow), and/or it might function downstream of incisions, during the HR step (not shown).
References
-
- Alpi AF, Pace PE, Babu MM, Patel KJ. Mechanistic insight into site-restricted monoubiquitination of FANCD2 by Ube2t, FANCL, and FANCI. Molecular cell. 2008;32:767–777. - PubMed
-
- Aravind L, Koonin EV. SAP - a putative DNA-binding motif involved in chromosomal organization. Trends Biochem Sci. 2000;25:112–114. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
