

Reconstruction of the mouse otocyst and early neuroblast lineage at single-cell resolution
- PMID: 24768691
- PMCID: PMC4051200
- DOI: 10.1016/j.cell.2014.03.036
Reconstruction of the mouse otocyst and early neuroblast lineage at single-cell resolution
Abstract
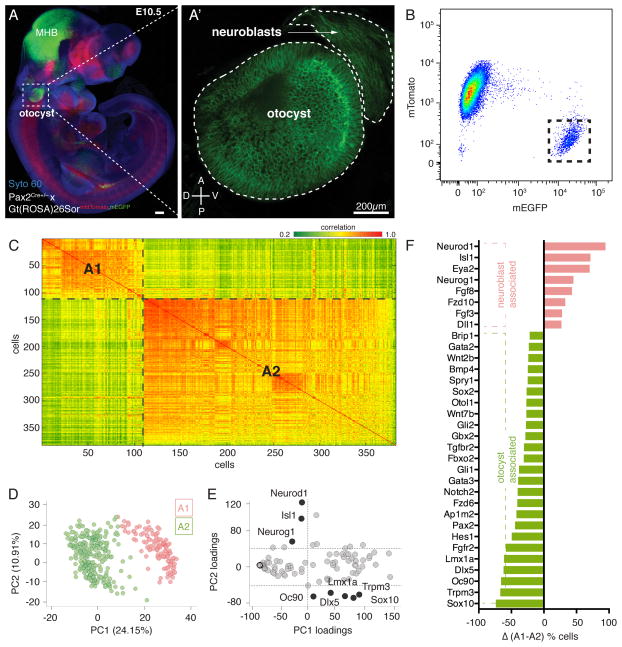
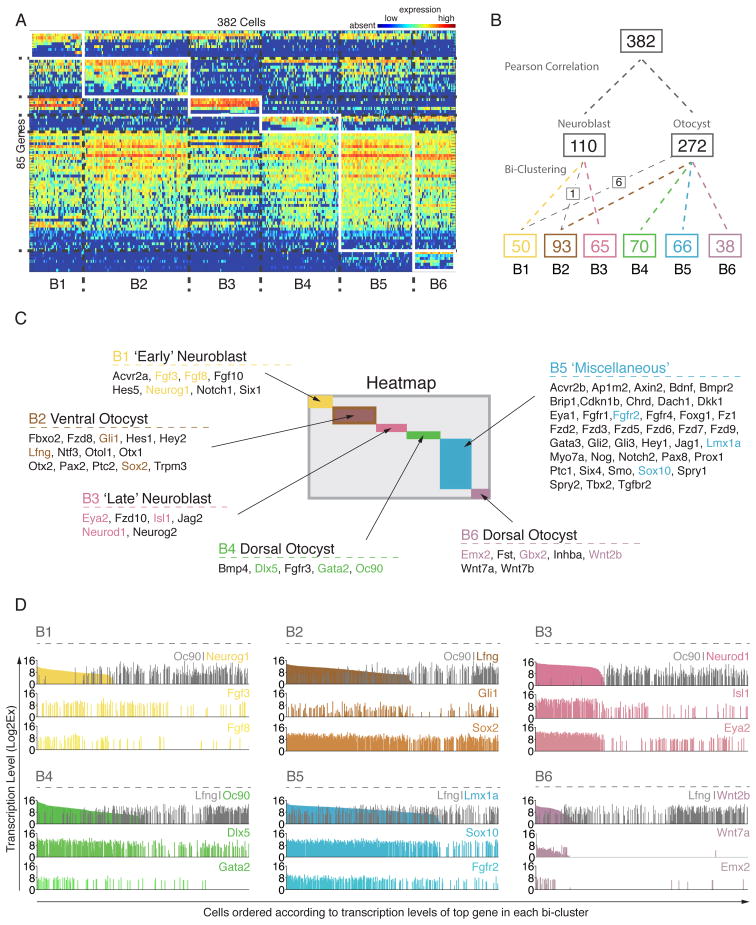
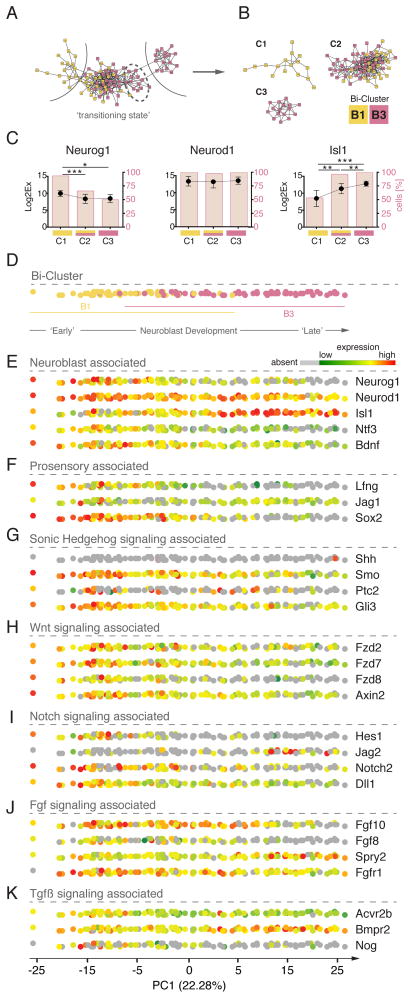
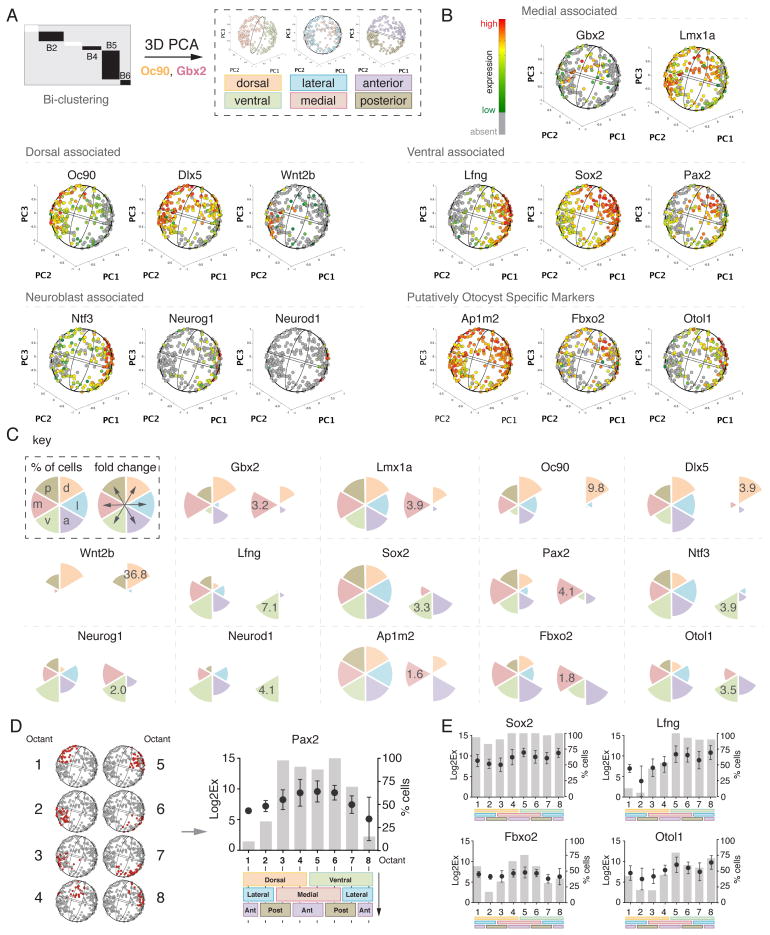
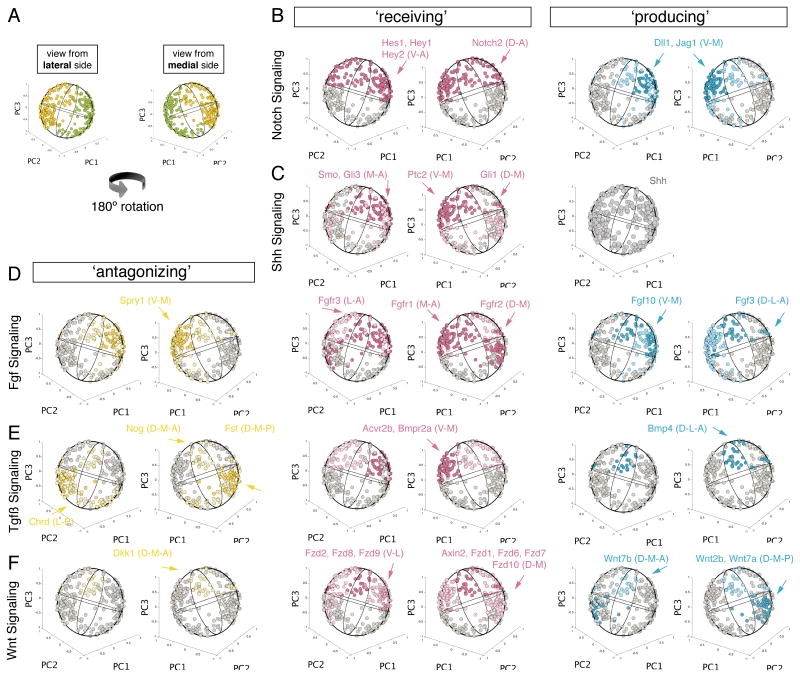
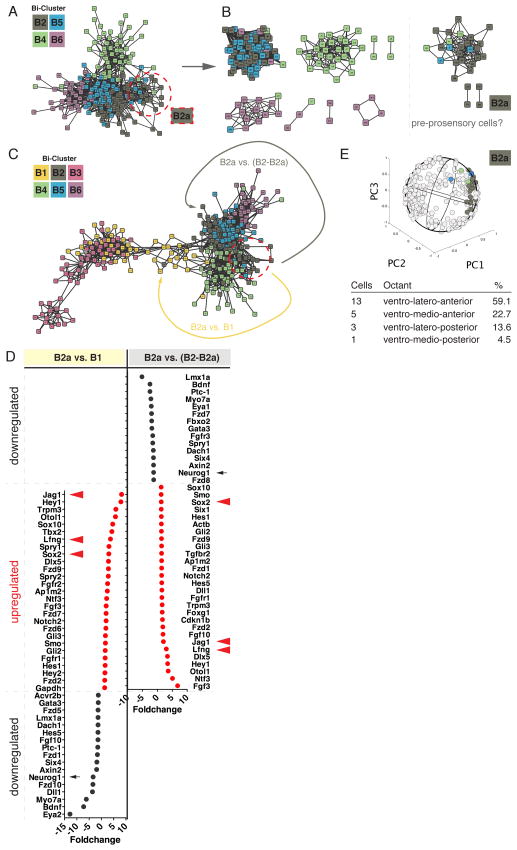
The otocyst harbors progenitors for most cell types of the mature inner ear. Developmental lineage analyses and gene expression studies suggest that distinct progenitor populations are compartmentalized to discrete axial domains in the early otocyst. Here, we conducted highly parallel quantitative RT-PCR measurements on 382 individual cells from the developing otocyst and neuroblast lineages to assay 96 genes representing established otic markers, signaling-pathway-associated transcripts, and novel otic-specific genes. By applying multivariate cluster, principal component, and network analyses to the data matrix, we were able to readily distinguish the delaminating neuroblasts and to describe progressive states of gene expression in this population at single-cell resolution. It further established a three-dimensional model of the otocyst in which each individual cell can be precisely mapped into spatial expression domains. Our bioinformatic modeling revealed spatial dynamics of different signaling pathways active during early neuroblast development and prosensory domain specification.
Copyright © 2014 Elsevier Inc. All rights reserved.
Figures
Comment in
-
Reconstructing complex tissues from single-cell analyses.Cell. 2014 May 8;157(4):771-3. doi: 10.1016/j.cell.2014.04.024. Cell. 2014. PMID: 24813603
References
-
- Abello G, Khatri S, Giraldez F, Alsina B. Early regionalization of the otic placode and its regulation by the Notch signaling pathway. Mech Dev. 2007;124:631–645. - PubMed
-
- Alsina B, Abello G, Ulloa E, Henrique D, Pujades C, Giraldez F. FGF signaling is required for determination of otic neuroblasts in the chick embryo. Dev Biol. 2004;267:119–134. - PubMed
-
- Alsina B, Giraldez F, Pujades C. Patterning and cell fate in ear development. Int J Dev Biol. 2009;53:1503–1513. - PubMed
-
- Bok J, Dolson DK, Hill P, Ruther U, Epstein DJ, Wu DK. Opposing gradients of Gli repressor and activators mediate Shh signaling along the dorsoventral axis of the inner ear. Development. 2007;134:1713–1722. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
