

Developmental studies of the hippocampus and hippocampal-dependent behaviors: insights from interdisciplinary studies and tips for new investigators
- PMID: 24769291
- PMCID: PMC4939092
- DOI: 10.1016/j.neubiorev.2014.04.009
Developmental studies of the hippocampus and hippocampal-dependent behaviors: insights from interdisciplinary studies and tips for new investigators
Abstract
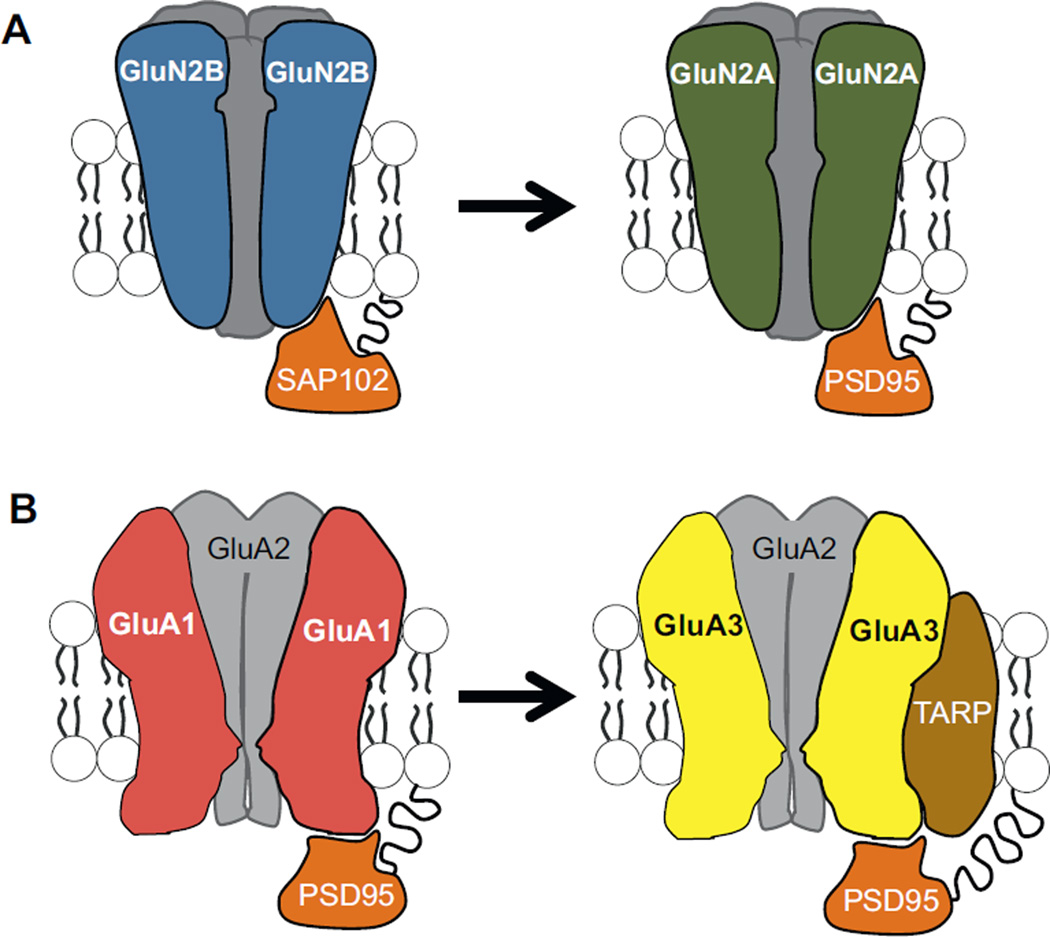
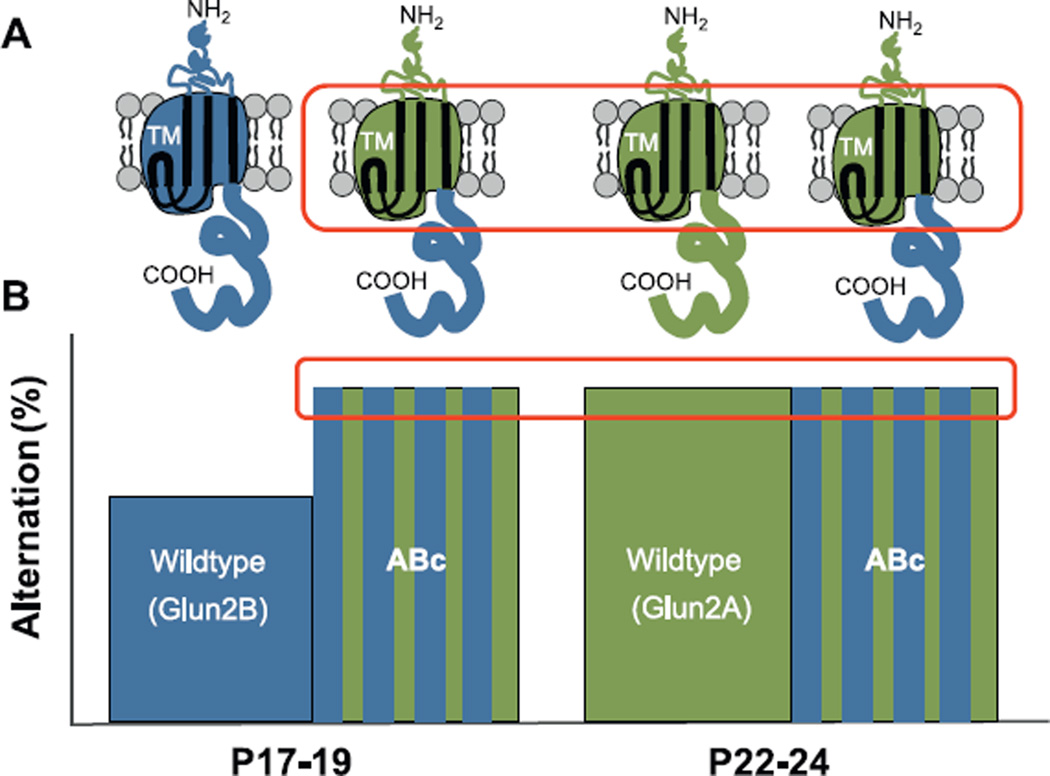
The hippocampus is not fully developed at birth and, with respect to spatial cognition, only begins to show signs of adult-like function at three postnatal weeks in rodents. Studying the developmental period spanning roughly two to four weeks of age permits an understanding of the neural framework necessary for the emergence of spatial navigation and, quite possibly, human episodic memory. However, due to developmental factors, behavior data collection and interpretation can be severely compromised if inappropriate designs are applied. As such, we propose methodological considerations for the behavioral assessment of hippocampal function in developing rats that take into account animal size, growth rate, and sensory and motor ability. We further summarize recent key interdisciplinary studies that are beginning to unravel the molecular machinery and physiological alterations responsible for hippocampal maturation. In general, hippocampal development is a protracted process during which unique contributions to spatial cognition and complex recognition memory come "on line" at different postnatal ages creating a unique situation for elucidating the neural bases of specific components of higher cognitive abilities.
Keywords: AMPAR; Behavioral testing; Environment; Hippocampus; Juvenile; Learning and memory; NMDAR; Novelty; Place field; Postnatal development; Spatial navigation; Y-maze.
Copyright © 2014 Elsevier Ltd. All rights reserved.
Figures
References
-
- Aadland J, Beatty WW, Maki RH. Spatial memory of children and adults assessed in the radial maze. Dev. Psychobiol. 1985;18:163–172. http://dx.doi.org/10.1002/dev.420180208. - DOI - PubMed
-
- Ainge JA, Tamosiunaite M, Woergoetter F, Dudchenko PA. Hippocampal CA1 place cells encode intended destination on a maze with multiple choice points. J. Neurosci. 2007;27:9769–9779. http://dx.doi.org/10.1523/JNEUROSCI.2011-07.2007. - DOI - PMC - PubMed
-
- Ainge JA, Tamosiunaite M, Worgotter F, Dudchenko PA. Hippocampal place cells encode intended destination, and not a discriminative stimulus, in a conditional T-maze task. Hippocampus. 2012;22:534–543. http://dx.doi.org/10.1002/hipo.20919. - DOI - PubMed
-
- Arai AC, Kessler M. Pharmacology of ampakine modulators: from AMPA receptors to synapses and behavior. Curr. Drug Targets. 2007;8:583–602. http://dx.doi.org/10.2174/138945007780618490. - DOI - PubMed
-
- Barker GR, Warburton EC. When is the hippocampus involved in recognition memory? J. Neurosci. 2011;31:10721–10731. http://dx.doi.org/10.1523/JNEUROSCI.6413-10.2011. - DOI - PMC - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
