

Methylcrotonyl-CoA Carboxylase Regulates Triacylglycerol Accumulation in the Model Diatom Phaeodactylum tricornutum
- PMID: 24769481
- PMCID: PMC4036579
- DOI: 10.1105/tpc.114.124982
Methylcrotonyl-CoA Carboxylase Regulates Triacylglycerol Accumulation in the Model Diatom Phaeodactylum tricornutum
Abstract
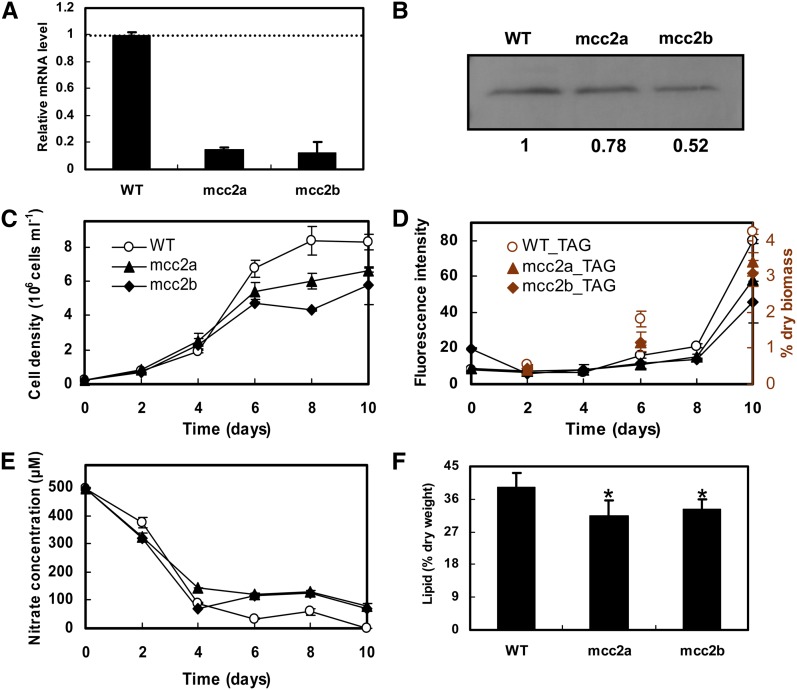
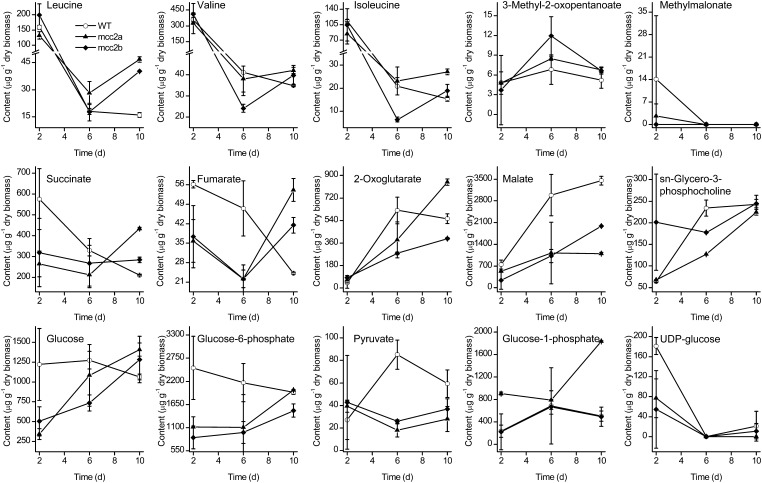
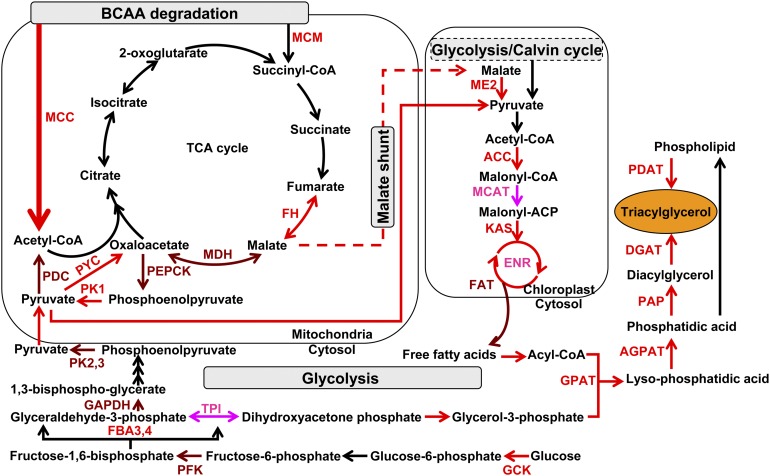
The model marine diatom Phaeodactylum tricornutum can accumulate high levels of triacylglycerols (TAGs) under nitrogen depletion and has attracted increasing attention as a potential system for biofuel production. However, the molecular mechanisms involved in TAG accumulation in diatoms are largely unknown. Here, we employed a label-free quantitative proteomics approach to estimate differences in protein abundance before and after TAG accumulation. We identified a total of 1193 proteins, 258 of which were significantly altered during TAG accumulation. Data analysis revealed major changes in proteins involved in branched-chain amino acid (BCAA) catabolic processes, glycolysis, and lipid metabolic processes. Subsequent quantitative RT-PCR and protein gel blot analysis confirmed that four genes associated with BCAA degradation were significantly upregulated at both the mRNA and protein levels during TAG accumulation. The most significantly upregulated gene, encoding the β-subunit of methylcrotonyl-CoA carboxylase (MCC2), was selected for further functional studies. Inhibition of MCC2 expression by RNA interference disturbed the flux of carbon (mainly in the form of leucine) toward BCAA degradation, resulting in decreased TAG accumulation. MCC2 inhibition also gave rise to incomplete utilization of nitrogen, thus lowering biomass during the stationary growth phase. These findings help elucidate the molecular and metabolic mechanisms leading to increased lipid production in diatoms.
© 2014 American Society of Plant Biologists. All rights reserved.
Figures
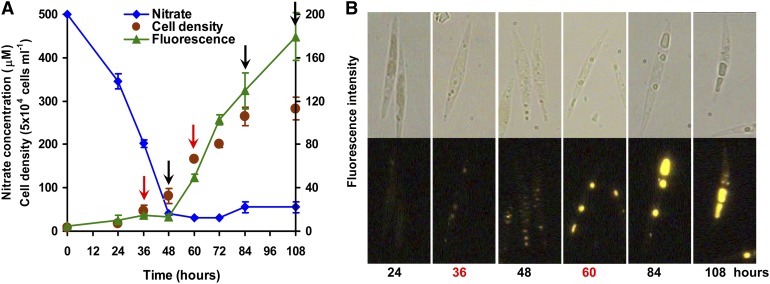
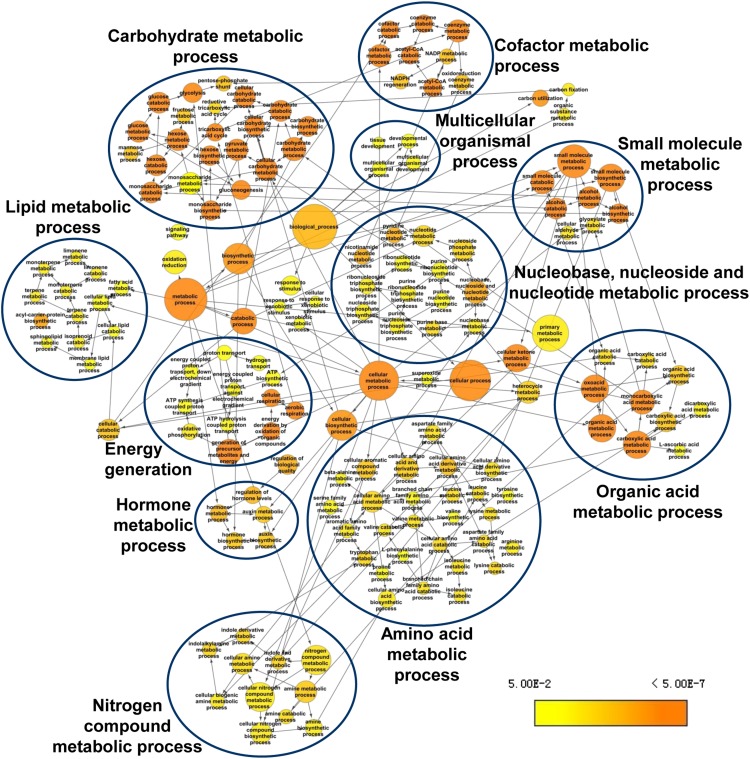
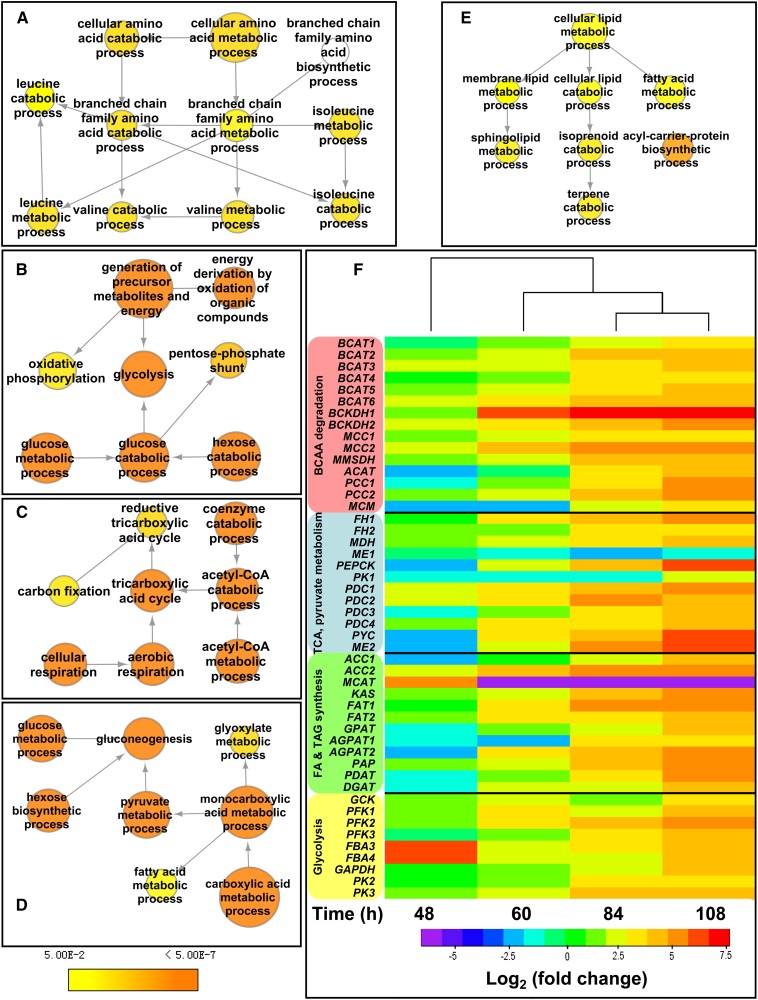
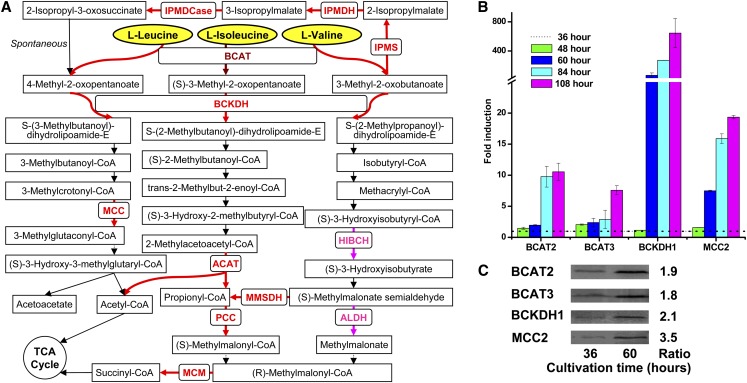
References
-
- Aguirre A.-M., Bassi A., Saxena P. (2013). Engineering challenges in biodiesel production from microalgae. Crit. Rev. Biotechnol. 33: 293–308 - PubMed
-
- Allen A.E., Dupont C.L., Oborník M., Horák A., Nunes-Nesi A., McCrow J.P., Zheng H., Johnson D.A., Hu H., Fernie A.R., Bowler C. (2011). Evolution and metabolic significance of the urea cycle in photosynthetic diatoms. Nature 473: 203–207 - PubMed
-
- Bantscheff M., Lemeer S., Savitski M.M., Kuster B. (2012). Quantitative mass spectrometry in proteomics: critical review update from 2007 to the present. Anal. Bioanal. Chem. 404: 939–965 - PubMed
-
- Bender S.J., Parker M.S., Armbrust E.V. (2012). Coupled effects of light and nitrogen source on the urea cycle and nitrogen metabolism over a diel cycle in the marine diatom Thalassiosira pseudonana. Protist 163: 232–251 - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
