

Identification of a ZEB2-MITF-ZEB1 transcriptional network that controls melanogenesis and melanoma progression
- PMID: 24769727
- PMCID: PMC4085532
- DOI: 10.1038/cdd.2014.44
Identification of a ZEB2-MITF-ZEB1 transcriptional network that controls melanogenesis and melanoma progression
Abstract
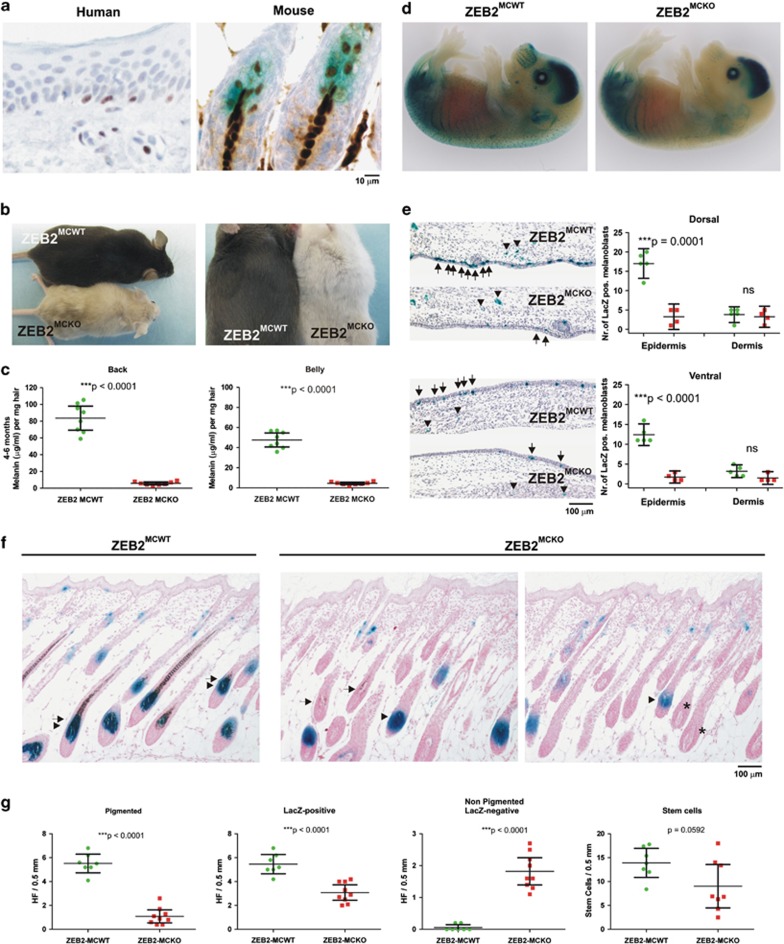
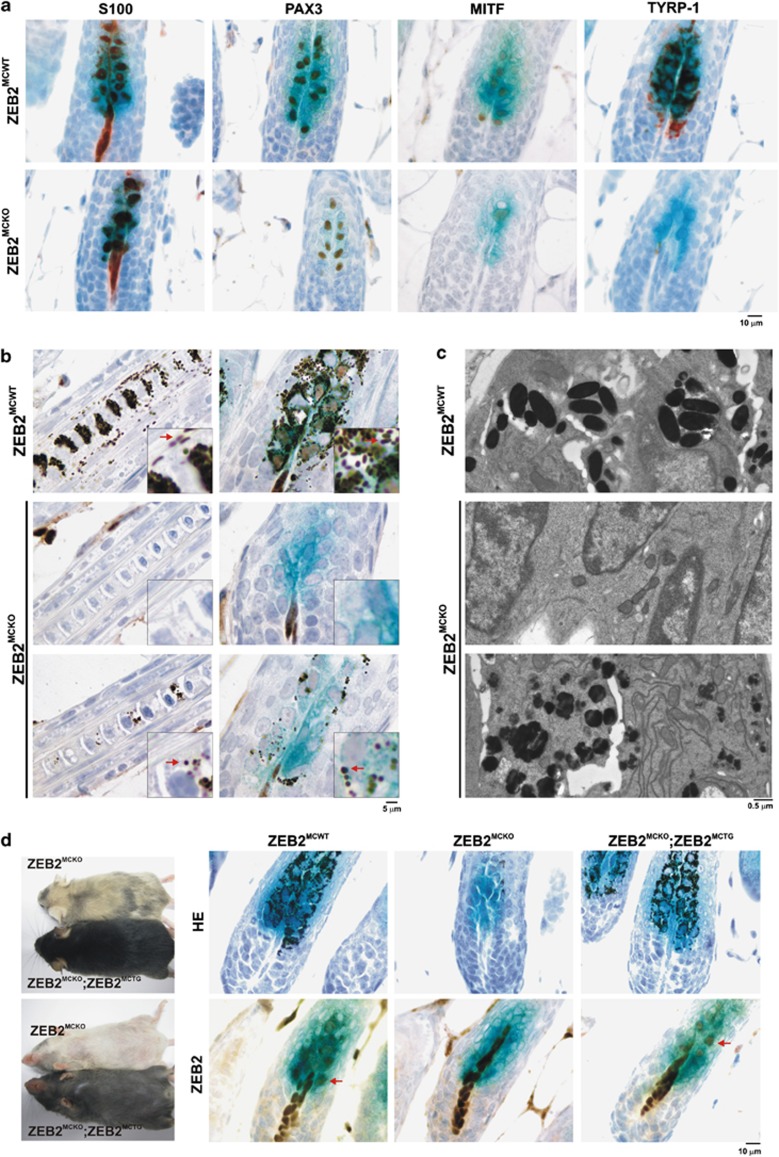
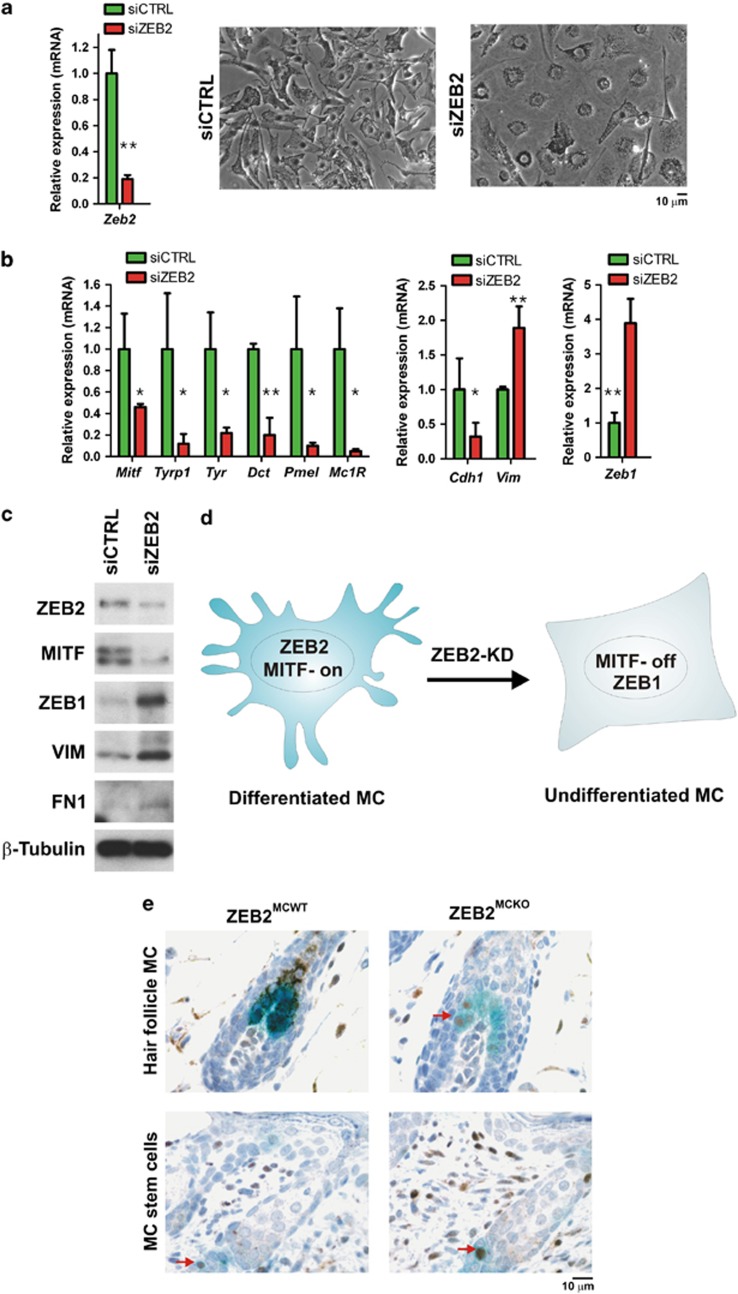
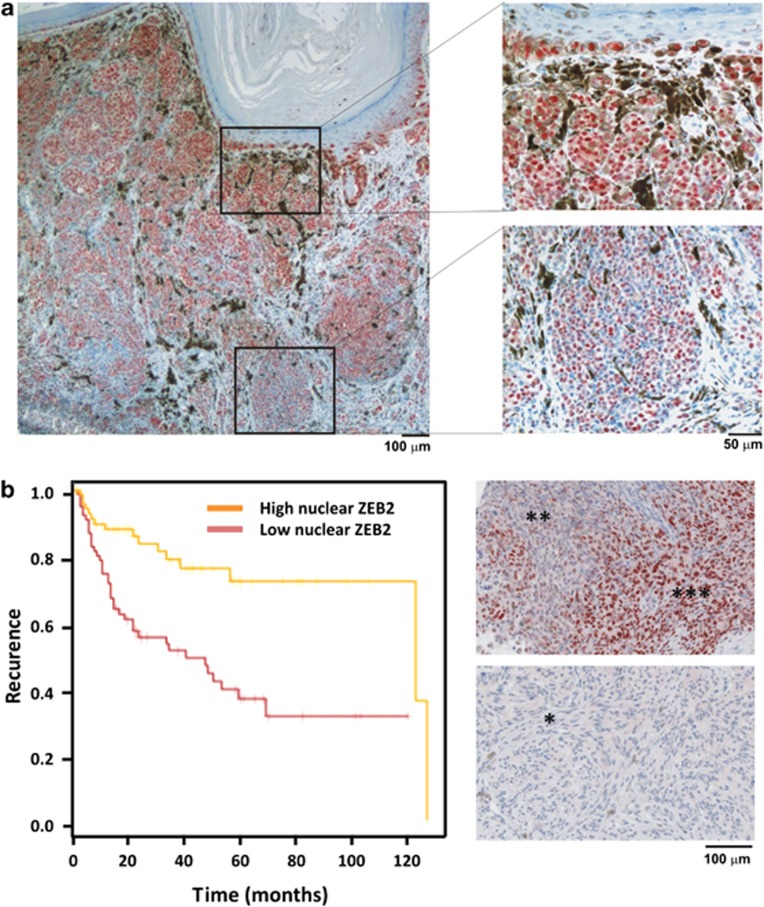
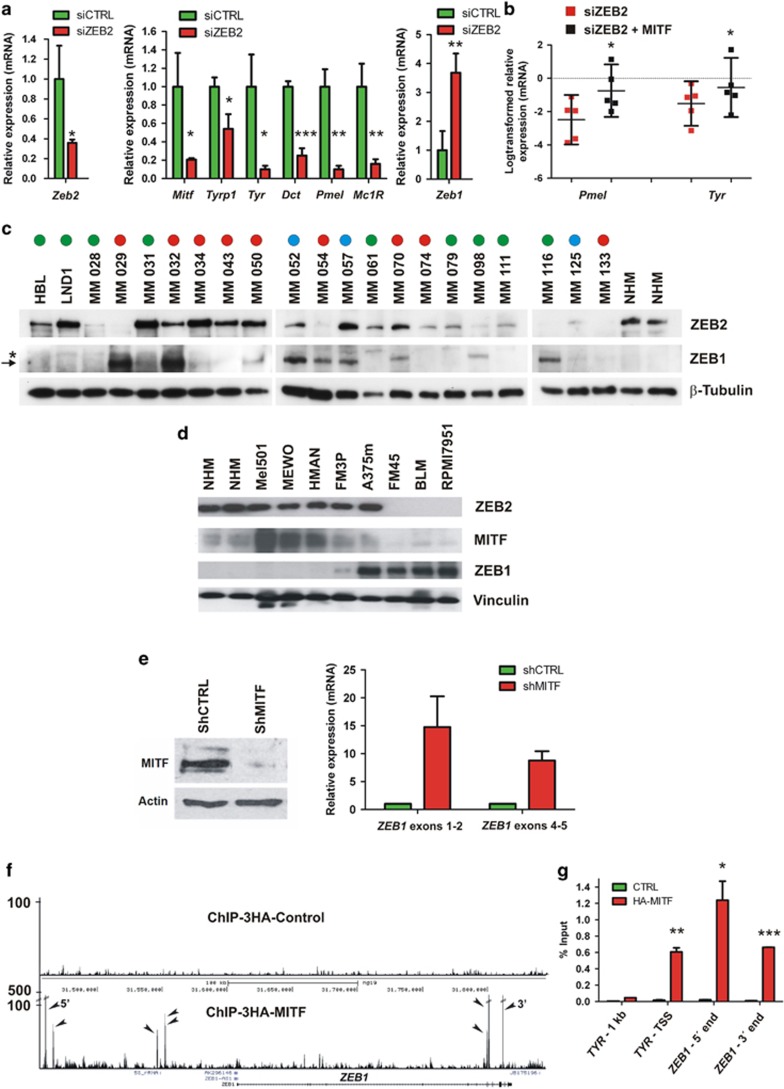
Deregulation of signaling pathways that control differentiation, expansion and migration of neural crest-derived melanoblasts during normal development contributes also to melanoma progression and metastasis. Although several epithelial-to-mesenchymal (EMT) transcription factors, such as zinc finger E-box binding protein 1 (ZEB1) and ZEB2, have been implicated in neural crest cell biology, little is known about their role in melanocyte homeostasis and melanoma. Here we show that mice lacking Zeb2 in the melanocyte lineage exhibit a melanoblast migration defect and, unexpectedly, a severe melanocyte differentiation defect. Loss of Zeb2 in the melanocyte lineage results in a downregulation of the Microphthalmia-associated transcription factor (Mitf) and melanocyte differentiation markers concomitant with an upregulation of Zeb1. We identify a transcriptional signaling network in which the EMT transcription factor ZEB2 regulates MITF levels to control melanocyte differentiation. Moreover, our data are also relevant for human melanomagenesis as loss of ZEB2 expression is associated with reduced patient survival.
Figures
References
-
- Ernfors P. Cellular origin and developmental mechanisms during the formation of skin melanocytes. Exp Cell Res. 2010;316:1397–1407. - PubMed
-
- Luciani F, Champeval D, Herbette A, Denat L, Aylaj B, Martinozzi S, et al. Biological and mathematical modeling of melanocyte development. Development. 2011;138:3943–3954. - PubMed
-
- Osawa M.Melanocyte stem cells(June 30, 2009), StemBook, ed. The Stem Cell Research Community, StemBook,doi: 10.3824/stembook.1.46.1 - DOI
-
- Steingrimsson E, Copeland NG, Jenkins NA. Melanocyte stem cell maintenance and hair graying. Cell. 2005;121:9–12. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
