

Chromatin retention of DNA damage sensors DDB2 and XPC through loss of p97 segregase causes genotoxicity
- PMID: 24770583
- PMCID: PMC4007632
- DOI: 10.1038/ncomms4695
Chromatin retention of DNA damage sensors DDB2 and XPC through loss of p97 segregase causes genotoxicity
Abstract
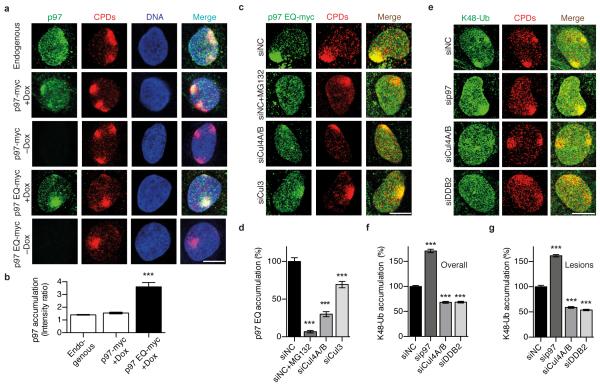
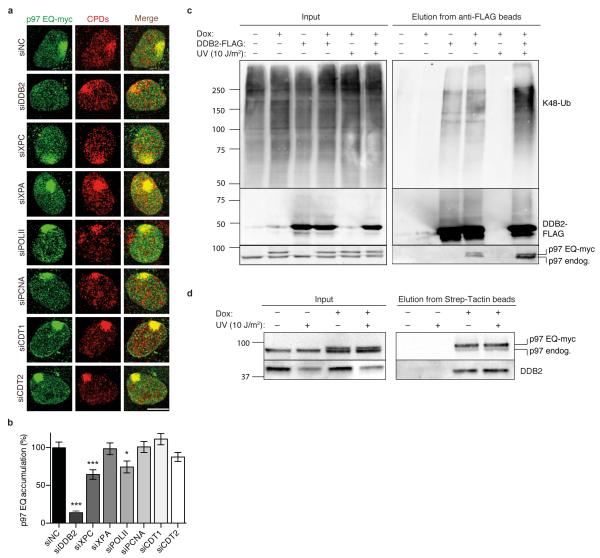
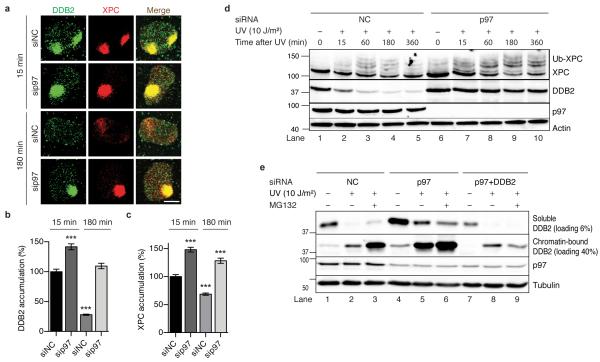
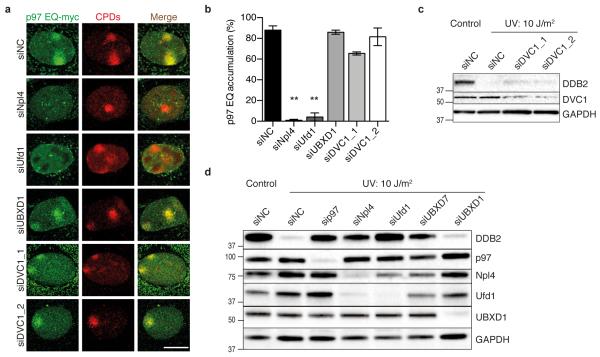
DNA damage recognition subunits such as DDB2 and XPC protect the human skin from ultraviolet (UV) light-induced genome instability and cancer, as demonstrated by the devastating inherited syndrome xeroderma pigmentosum. Here we show that the beneficial DNA repair response triggered by these two genome caretakers critically depends on a dynamic spatiotemporal regulation of their homeostasis. The prolonged retention of DDB2 and XPC in chromatin, because of a failure to readily remove both recognition subunits by the ubiquitin-dependent p97/VCP/Cdc48 segregase complex, leads to impaired DNA excision repair of UV lesions. Surprisingly, the ensuing chromosomal aberrations in p97-deficient cells are alleviated by a concomitant downregulation of DDB2 or XPC. Also, genome instability resulting from an excess of DDB2 persisting in UV-irradiated cells is prevented by concurrent p97 overexpression. Our findings demonstrate that DNA damage sensors and repair initiators acquire unexpected genotoxic properties if not controlled by timely extraction from chromatin.
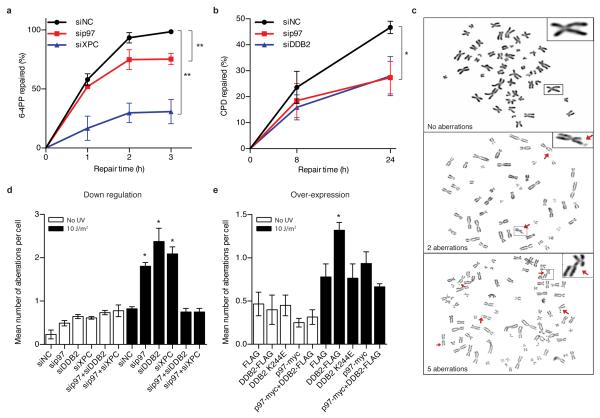
Figures
Similar articles
-
Ubiquitin-specific protease 7 regulates nucleotide excision repair through deubiquitinating XPC protein and preventing XPC protein from undergoing ultraviolet light-induced and VCP/p97 protein-regulated proteolysis.J Biol Chem. 2014 Sep 26;289(39):27278-27289. doi: 10.1074/jbc.M114.589812. Epub 2014 Aug 12. J Biol Chem. 2014. PMID: 25118285 Free PMC article.
-
Cullin 4A-mediated proteolysis of DDB2 protein at DNA damage sites regulates in vivo lesion recognition by XPC.J Biol Chem. 2006 May 12;281(19):13404-13411. doi: 10.1074/jbc.M511834200. Epub 2006 Mar 8. J Biol Chem. 2006. PMID: 16527807
-
DDB2 promotes chromatin decondensation at UV-induced DNA damage.J Cell Biol. 2012 Apr 16;197(2):267-81. doi: 10.1083/jcb.201106074. Epub 2012 Apr 9. J Cell Biol. 2012. PMID: 22492724 Free PMC article.
-
The role of ubiquitin-dependent segregase p97 (VCP or Cdc48) in chromatin dynamics after DNA double strand breaks.Philos Trans R Soc Lond B Biol Sci. 2017 Oct 5;372(1731):20160282. doi: 10.1098/rstb.2016.0282. Philos Trans R Soc Lond B Biol Sci. 2017. PMID: 28847819 Free PMC article. Review.
-
Molecular mechanisms of DNA damage recognition for mammalian nucleotide excision repair.DNA Repair (Amst). 2016 Aug;44:110-117. doi: 10.1016/j.dnarep.2016.05.015. Epub 2016 May 20. DNA Repair (Amst). 2016. PMID: 27264556 Review.
Cited by
-
Dissecting regulatory pathways for transcription recovery following DNA damage reveals a non-canonical function of the histone chaperone HIRA.Nat Commun. 2021 Jun 22;12(1):3835. doi: 10.1038/s41467-021-24153-1. Nat Commun. 2021. PMID: 34158510 Free PMC article.
-
Timing of DNA lesion recognition: Ubiquitin signaling in the NER pathway.Cell Cycle. 2017 Jan 17;16(2):163-171. doi: 10.1080/15384101.2016.1261227. Epub 2016 Dec 8. Cell Cycle. 2017. PMID: 27929739 Free PMC article. Review.
-
Structure and function of the AAA+ ATPase p97/Cdc48p.Gene. 2016 May 25;583(1):64-77. doi: 10.1016/j.gene.2016.02.042. Epub 2016 Mar 3. Gene. 2016. PMID: 26945625 Free PMC article. Review.
-
Pharmacological targeting of valosin containing protein (VCP) induces DNA damage and selectively kills canine lymphoma cells.BMC Cancer. 2015 Jun 24;15:479. doi: 10.1186/s12885-015-1489-1. BMC Cancer. 2015. PMID: 26104798 Free PMC article.
-
ROS-Induced Activation of DNA Damage Responses Drives Senescence-Like State in Postmitotic Cochlear Cells: Implication for Hearing Preservation.Mol Neurobiol. 2019 Aug;56(8):5950-5969. doi: 10.1007/s12035-019-1493-6. Epub 2019 Jan 28. Mol Neurobiol. 2019. PMID: 30693443 Free PMC article.
References
-
- Friedberg EC, et al. DNA repair: from molecular mechanism to human disease. DNA Repair (Amst) 2006;5:986–996. - PubMed
-
- Hoeijmakers JH. DNA damage, aging, and cancer. N. Engl. J. Med. 2009;361:1475–1485. - PubMed
-
- Gillet LC, Scharer OD. Molecular mechanisms of mammalian global genome nucleotide excision repair. Chem. Rev. 2006;106:253–276. - PubMed
-
- Bergink S, Jaspers NG, Vermeulen W. Regulation of UV-induced DNA damage response by ubiquitylation. DNA Rep. (Amst) 2007;6:1231–1242. - PubMed
-
- Hanawalt PC, Spivak G. Transcription-coupled DNA repair: two decades of progress and surprises. Nat. Rev. Mol. Cell Biol. 2008;9:958–970. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
